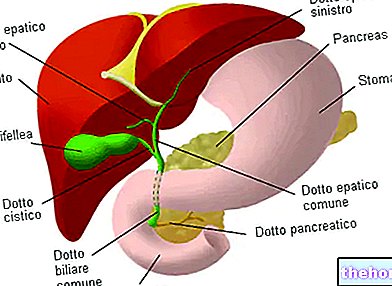
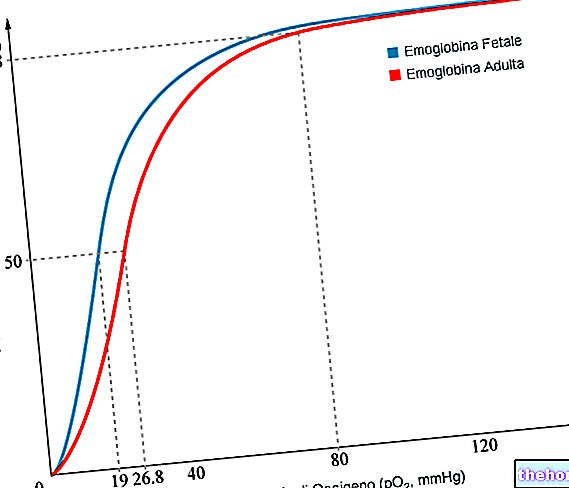
発生学の注記
発生学は、接合子からそのすべての器官とシステムに恵まれた生物までの一連の発達形態を研究します。
この点で、とりわけ定量的な意味で意図された、開発(複雑さが増す構造的および組織的フェーズの継承)と成長の違いを覚えておく価値があります。
私たちが目撃する無脊椎動物の後生動物では、(円口類、魚類、両生類、爬虫類、鳥類、哺乳類を介して)人間に至るまでの進化のシリーズで、複雑さが増す成体の出現が見られます。
最初に、常に予備材料を備えた接合子は、種の正常な核/細胞質比に達するまで、成長せずに割球と呼ばれる2、次に4、次に8などの細胞に(その後の有糸分裂によって)分割されます。
この最初のセグメンテーションは、重水素の量と分布に応じて、さまざまなパターンに従うことができます。
当初、重水素は希少であるため(「卵黄卵黄」)、セグメンテーションは完全であり、非常に異なる割球を生じます。胚の複雑さが増すにつれて、その発達が始まるまでに、より多くの時間と材料が必要になります。独立した生活。これには、接合子の一部に配置される傾向があるdeutoplasma(「telolecyticeggs」)の増加が必要です。これにより、セグメンテーションの変更に関連する「異方性」が増加します。これは、次の2つの一般原則によって管理されます。
-Hertwigの法則によれば、有糸分裂では、無彩色の紡錘体(赤道が娘細胞の分裂面を決定する)は細胞質の最大長の方向に配置される傾向があります。
-バルフォアの法則によれば、セグメンテーション速度は重水素の量に反比例します。
円口類と魚ではすでに分節化が不均等であり、急速に分節化された動物の極(胚の上部構造を与える)と卵黄の極が予備物質の大部分を含んでいることがわかります。この傾向はさらに大きくなります。 。卵黄極がゆっくりと分節している間、比較的不活性のままであり、急速に分節している動物の極に由来する細胞によって覆われることになる、両生類(空気呼吸に関与する器官を準備する必要がある)の異方性。主な胚の段階には、接合子、割球、モルラ(ブラックベリーに似た割球のクラスター)、ブラストラ(内部細胞が退行したモルラ)、ガストルラ(片側の細胞が陥入したブラストラ)が含まれます。外部細胞層(神経系が最初に派生する外胚葉)とintを備えた生物のエルノ(内胚葉)、その間に第3層(中胚葉)が介在します。これらの層または「胚シート」から、順序付けられた順序で、すべての組織、臓器、およびシステムが導き出されます。
さらに進化した種では、deutoplasm(または「子牛」)の増加はそれがセグメント化することさえできないようなものです。したがって、鳥では、セグメント化は薄い表面の円盤にのみ影響し、「discoblastula」と一連の上記とは異なる方法で胚の形成を保証する現象。
卵黄のさらなる増加はおそらくより効率的ではなかったので、哺乳類では、独立した生活の能力までの発達と成長が別のシステムで得られます。実際、哺乳類では、卵黄は最初の段階でのみ使用されることに注意してください。その後、胚は母体との代謝関係を確立し(胎盤の出現を伴う)、過剰な卵黄を使用しなくなります。この時点で、卵黄は卵黄に戻り、セグメンテーションを元に戻すことができます。完全である(したがって、初期段階では「両生類」のそれと似ています)が、モルラの後、胚形成は鳥の最も進化したパターンに従って継続し、「芽球嚢胞」に続いて子宮壁に移植されます。そのため、胚の代謝は、卵黄ではなく母体によって(胎盤を介して)確保されます。
胚の分化
接合子のセグメンテーションが核/細胞質比を種の標準にもたらしたとき、成長も発達と並行して開始しなければなりません。このため、核小体とタンパク質合成の出現とともに代謝が始まります。このように開始されたタンパク質合成は、胚発生の初期段階に関与する遺伝子によるものです。これらの遺伝子は、動物と子牛の極のさまざまな割球に存在する物質によって抑制されます。次に、これらの初期遺伝子の産物は、後期段階を担当する遺伝子のオペロンを抑制解除することができます。この第2の一連の遺伝子の産物は、新しい胚構造を構築するという意味と、前のオペロンを抑制し、次のオペロンを抑制解除するという意味の両方で、新しい生物の構築につながる順序付けられた順序で作用することができます。 、蓄積された遺伝子情報のおかげで、ゲノムから数千年に渡って、さらに進化した種になりました。
ヘッケルの有名な表現「個体発生は系統発生を要約する」は、実際には、高等種が胚発生の段階で、進化的に前の種ですでに見られた継承を繰り返すという事実を表現しています。
胚の初期段階は、特に鰓の出現まで、脊椎動物で類似している傾向があります。
空中呼吸に切り替わる種では、鰓は再吸収されて再利用されますが(たとえば内分泌腺の形成のため)、鰓の形成に関連する遺伝情報は人間にも保存されます。これは明らかに、すべての脊椎動物のゲノムに存在し、個体発生の瞬間に機能した後も抑制されたままでなければならない胚の構造遺伝子の例です。
遺伝子作用の調節という意味での胚発生の解釈は、実験発生学の複雑な伝統的な経験を統合することを可能にします。
双子
接合子と最初の割球は、タンパク質合成が始まるまで、全能性です。つまり、生物全体に生命を与えることができます。これには、両生類の接合子の絞扼から2つの胚を得たシュペーマンの実験が関係しています。同様の現象は、人間の同一の双子の現象に基づいて現れます。これは、まさにこの理由で一卵性双生児(MZ)と呼ばれます。Spemannの実験的な双子は正常の半分のサイズでしたが、人間では完全に正常です。これは、両生類では2つの胚がすでに受け取った唯一の卵黄を共有する必要があったのに対し、男性では胎盤を介して胚の発達と成長に必要なすべてのものを受け取ることができるためです。
「双子の場合の3分の2の人には」別の起源があることを覚えておく必要があります。それらは、受精すると2つの接合子を与える2つの胚珠の放出を伴う、2つの卵胞の時折の同時成熟に由来します。この場合、私たちは二卵性双生児(DZ)について話します。
単一の接合子からの有糸分裂によって分割されたMZ双子は同じゲノムを持っているので、それらの間の違いは環境起源であるに違いありません。人間の遺伝学やスポーツの分野でも広く使用されています。
「特定の倫理的理由により実験が禁止される男性では、遺伝的要因によってどのような性格が規制されているかを確認できます。実際、厳密に継承された性格(血液型など)は常にMZ双子でのみ一致します。 MZの形質の一致はDZの形質の一致に近く、その表現型の形質を決定する際には、環境要因が遺伝的要因よりも優勢であると推定されます。