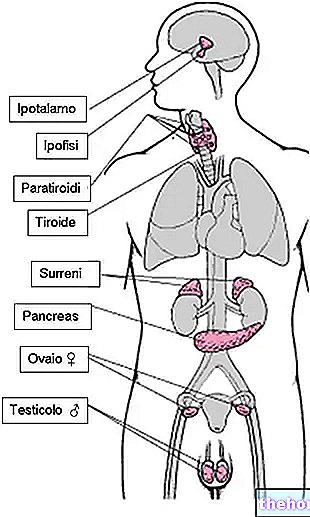
参照:ケトン食療法;糖尿病性ケトアシドーシス。
一般性
過去には、ケトン体は過剰な脂肪や糖尿病の摂取によって引き起こされた過剰な代謝によるものと考えられていましたが、一方、ケトン体は私たちの体によって自然に生成されます:脳はこれらの代謝物を条件で使用するように適応します長時間の絶食(糖尿病患者では、ケトン体がグルコース代謝に取って代わります)さらに、栄養不良の場合、ケトン体の経路が悪化する可能性があります。
ケトン体とは何ですか
ケトン体は脂質の誘導体です(脂質の代謝に由来し、ほとんどが肝臓に由来します)が、糖に似た特徴があります。
- 高い入力速度;
- すぐに使用できます。
一部のアミノ酸、特に代謝条件でさえ、ケトン体(ロイシン、リジン、フェニルアラニン、イソロイシン、トリプトファン、およびチロシン)を生成する可能性があります。
生物学的役割
- ケトン体はサイズが小さいため、非常に迅速に輸送されます(一方、アルブミンなどの輸送タンパク質を必要とする脂肪酸よりもはるかに多く)。
- ケトン体は、ほぼ独占的に筋肉と末梢組織によって使用されますが、心臓(使用するエネルギーの20〜30%はケトン体から供給されます)と脳(長時間の絶食の場合)によっても使用されます。
合成
ケトン体は、脂肪酸の代謝に由来するアセチル補酵素Aによって合成されます。
最初の段階を触媒する酵素は Β-ケトチオラーゼ、アセチル補酵素Aの硫黄を利用して、β-ケトアシル補酵素Aを生成します(これは、脂肪酸のβ酸化で見られる反応とは逆の反応です)。この反応は自発的ではなく、その後の反応によって引き起こされます。 、から触媒ヒドロキシメチルグルタリル補酵素Aシンターゼ これは、第2のアセチル補酵素Aの結合を含み、3-ヒドロキシ3-メチルグルタリル補酵素Aを取得します。
続いて、溶解酵素が介入し、3-ヒドロキシ3-メチルグルタリル補酵素Aをケトン体である酢酸酢に変換します。酢酸酢は末梢組織に送ることができます、または酵素の作用によって ヒドロキシ酪酸デヒドロゲナーゼ、3-β-ヒドロキシブチレートに変換。酢酸酢が非常に高濃度である場合、それはまた自発的に脱炭酸してアセトンになる可能性があります。
アセトン、酢酸酢、3-β-ヒドロキシ酪酸は、私たちが考える3つのケトン体です。アセトンは、ケトン体の経路でランダムに生成され、呼気と蒸散によって排出される廃棄物です。
末梢組織での使用
肝臓で生成されたケトン体は、末梢組織に送られます。
酢酸酢と3-β-ヒドロキシ酪酸が末梢組織に到達するとどうなるか見てみましょう。酢酸酢はβ-ケト酸であるため、活性化されると、β酸化プロセスで使用して生成することができます。アセチル補酵素A:したがって、β-ケト酸をβ-ケトアシル補酵素Aに変換する必要があります。
酢酸酢が末梢組織の細胞のミトコンドリアに到達すると、酵素の作用を受けます。 スクシニル補酵素Aトランスフェラーゼ:この酵素を介して、酢酸酢はスクシニル補酵素A(クレブス回路に由来)と反応し、コハク酸塩と酢アセチル補酵素Aが得られます。
スクシニル補酵素Aを利用して酢酸酢を活性化することにより、GTPを生成する段階であるクレブス回路にジャンプします。これは、エネルギーの観点から、細胞がアセチル酢補酵素Aを取得するために喜んで支払うプロセスです。 ;後者はその後、 Β-ケトチオラーゼ (β酸化酵素)2分子のアセチル補酵素Aを生成し、クレブス回路に送ります。
3-β-ヒドロキシ酪酸が末梢組織に送られると、ミトコンドリア内の後者は、β-ヒドロキシ酪酸デヒドロゲナーゼ酵素の作用によってビネガーアセトンに変換され、約2.5ATPに相当するNADHが生成されます。生成された酢酸ビネガーは、前述の経路に従います。
末梢組織の細胞は、酢酸酢からではなく、3-β-ヒドロキシ酪酸からより多くのエネルギーを引き出しますが、末梢組織へのどちらか一方の送達は、肝臓のエネルギーの利用可能性に依存します。
C "は無視できない量の代謝脂肪酸であり、ミトコンドリアには含まれず、ペルオキシソームに含まれます。ペルオキシソームはミトコンドリアよりも小さく、金属イオンとペルオキシダーゼ酵素が豊富な細胞小器官です。ペルオキシダーゼ酵素は過酸化水素を使用してレドックスプロセスを促進します。は、過酸化水素を生成できる酵素システムです。
ペルオキシソームのβ酸化では、「アシル補酵素Aは」の作用によって得られます。アシル補酵素Aオキシダーゼ (ミトコンドリアでは、一方、酵素アシル補酵素Aデヒドロゲナーゼが作用しました)この場合も、トランス2,3エノイル補酵素Aが形成され、二機能性酵素の作用を受けます(それはと同じ機能を果たします) 「エノイル補酵素AヒドラターゼおよびL-β-ヒドロキシアシル補酵素Aデヒドロゲナーゼ)によってミトコンドリア内で生成され、したがって、β-ケトアシル補酵素Aに変換されます。この最後は、ミトコンドリアと同様に、β-ケトチオラーゼおよびアセチル補酵素Aの作用を受けます。アシル補酵素Aは、炭素質の骨格が最初の骨格と比較して2単位減少した状態で得られ、循環に戻ります。