
タンパク質の元素分析では、次の平均値が得られます:炭素55%、水素7%、窒素16%;タンパク質が互いに異なることは明らかですが、それらの平均元素組成は上記の値とほとんど異なりません。
構成上、タンパク質は天然のα-アミノ酸から形成された高分子です。アミノ酸は、α-アミノ酸のアミノ基と別のα-アミノ酸のカルボキシルとの間の反応によって確立されるアミド結合を介して結合します。
この結合(-CO-NH-)は、ペプチド(アミノ酸の組み合わせ)に結合するため、ペプチド結合とも呼ばれます。
得られたものは、2つのアミノ酸で構成されているため、ジペプチドです。ジペプチドは、一端に遊離アミノ基(NH2)、他端にカルボキシル基(COOH)を含むため、1つまたは複数のアミノ酸と反応し、同じ反応で右と左の両方から鎖を長くすることができます。上に見られる。
一連の反応(ちなみに、実際にはそれほど単純ではありません)は無期限に続く可能性があります:と呼ばれるポリマーが存在するまで ポリペプチド また タンパク質。ペプチドとタンパク質の違いは分子量に関連しています。通常、分子量が10,000を超える場合は、タンパク質について説明します。
最近、アミノ酸からタンパク質を自動的に生成する方法が開発され、優れた結果が得られていますが、アミノ酸を結合して小さなタンパク質を取得することは困難な作業です。
したがって、最も単純なタンパク質は2つのアミノ酸で構成されています。国際的な慣例により、タンパク質構造内のアミノ酸の順序付けられた番号付けは、遊離のα-アミノ基を持つアミノ酸から始まります。
無視できない化学的困難に遭遇するこのタンパク質のコーディング)。
エドマン分解によってアミノ酸の順序付けられた配列を決定することが可能でした。タンパク質はフェニルイソチオシアネート(FITC)と反応します。最初に、α-アミノ窒素ダブレットがフェニルイソチオシアネートを攻撃してチオカルバミル誘導体を形成します。続いて、得られた生成物は環化して、蛍光性であるフェニルチオヒダントイン誘導体を与える。
エドマンは、分解のパラメーター(時間、試薬、pHなど)を自動的に調整し、タンパク質の一次構造を提供するシーケンサーと呼ばれる機械を考案しました(このために彼はノーベル賞を受賞しました)。
一次構造は、タンパク質分子の特性を完全に解釈するのに十分ではありません。これらの特性は、本質的に、タンパク質分子が想定する傾向のある空間構成に依存し、さまざまな方法で折りたたまれると考えられています。つまり、タンパク質の二次構造として定義されているものを想定しています。
タンパク質の二次構造はちらつきます。つまり、加熱すると崩壊する傾向があります。その後、タンパク質は変性し、その特徴的な特性の多くを失います。 70°C以上に加熱することに加えて、変性は照射または反応物の作用(例えば強酸から)によっても引き起こされる可能性があります。
熱効果によるタンパク質の変性は、例えば卵白を加熱することによって観察されます:それはそのゼラチン状の外観を失い、不溶性の白い物質に変わるのが見られます。ただし、タンパク質の変性は二次構造の破壊につながりますが、一次構造は変更されません(さまざまなアミノ酸の連結)。
タンパク質は、その鎖が二次構造の曲がりにもかかわらずまだ柔軟であるが、固体の形でゆがんだ三次元配列を生み出すように折りたたまれるとき、三次構造をとる。三次構造の原因は、とりわけ、分子に沿って散在するシステイン-SH間に確立できるジスルフィド結合です。
一方、四次構造は、2つ以上のサブユニットによって形成されるタンパク質にのみ属します。たとえば、ヘモグロビンは、四面体の頂点に配置された2対のタンパク質(つまり、4つのタンパク質鎖すべて)で構成され、球形の構造を生成します。4つのタンパク質鎖は、イオン力とイオン力によって結合されます。非共有結合。
四次構造の別の例はインスリンの構造であり、中心に2つの亜鉛原子がある三角形の頂点にペアで配置された6つのタンパク質サブユニットで構成されているように見えます。
繊維状タンパク質
それらは一定の剛性を持ち、他のタンパク質よりもはるかに長い軸を持っているタンパク質です。自然界に大量に存在する繊維状タンパク質はコラーゲン(またはコラーゲン)です。
繊維状タンパク質は、さまざまな二次構造をとることができます。α-ヘリックス、β-シート、そしてコラーゲンの場合はトリプルヘリックス。 αヘリックスが最も安定した構造であり、次にβシートが続きますが、3つの中で最も安定性が低いのは三重らせんです。
右 主骨格(下から上に向けられている)に続いて、右側のねじのねじ込みと同様の動きが実行された場合。らせんが 左手 動きが左ねじのねじ込みに類似している場合右側のα-ヘリックスでは、アミノ酸の-R置換基はタンパク質の主軸に垂直であり、外側を向いていますが、左側では-手a-ヘリックス-R置換基は内側を向いています。右巻きのa-ヘリックスは左巻きのa-ヘリックスよりも安定しています。これは、vati -R c "の間で相互作用が少なく、立体障害が少ないためです。タンパク質に含まれるすべてのa-ヘリックスは右旋性です。
αヘリックスの構造は、各アミノ酸のカルボキシル基(-C = O)と4残基後に見つかったアミノ基(-NH)の間に形成される水素結合(水素結合)によって安定化されます線形シーケンス。
αヘリックス構造を有するタンパク質の例は、ヘアケラチンである。
αヘリックス構造を長くすることにより、αヘリックスからβシートへの移行が行われます。また、熱または機械的応力により、α-ヘリックスからβ-シート構造に移行することができます。
通常、タンパク質では、タンパク質自体の一部の間に鎖間水素結合を確立できるため、βシート構造は互いに近接しています。
繊維状タンパク質では、タンパク質構造のほとんどがα-ヘリックスまたはβ-シートで構成されています。
球状タンパク質
それらはほぼ球形の空間構造を持っています(ポリペプチド鎖の方向の多数の変化のため)。存在の一部はα-ヘリックスまたはβ-シート構造にまでさかのぼることができ、他の部分は代わりにこれらの形態に起因するものではありません。配置はランダムではなく、組織化され、反復的です。
これまで言及されてきたタンパク質は、完全に均質な構成の物質です。つまり、組み合わされたアミノ酸の純粋な配列です。そのようなタンパク質は言われています 単純;たんぱく質と呼ばれるたんぱく質部分と非たんぱく質部分(前立腺群)からなるたんぱく質があります 共役.
、爪、角膜、目の水晶体、一部の臓器(肝臓など)の間質腔の間など。
その構造はそれに特定の機械的能力を与えます。それは、それが実行しなければならない機能に応じて、高い弾性(例えば、腱)または高い剛性(例えば、骨)に関連する優れた機械的強度を有する。
コラーゲンの最も興味深い特性の1つは、その構成の単純さです。コラーゲンは、約30%のプロリンと約30%のグリシンで構成されています。他の18アミノ酸は、タンパク質構造の残りの40%を共有するだけで済みます。コラーゲンのアミノ酸配列は非常に規則的です。3残基ごとに3番目はグリシンです。
プロリンは、R基がα-アミノ窒素に結合する環状アミノ酸であり、これにより一定の剛性が得られます。
最終的な構造は、「らせん」の形をした反復鎖です。コラーゲン鎖内では、水素結合は存在しません。コラーゲンは「α-ヘリックスよりも大きいピッチ(ヘリックスの1回転に対応する長さ)を持つ左側のヘリックスです;コラーゲンヘリックスは非常に緩いので、3つのタンパク質鎖が互いに巻き付いて「」を形成することができますシングルロープ:トリプルヘリックス構造。
しかし、コラーゲンの三重らせんは、α-らせんとβ-シート構造の両方よりも安定性が低くなっています。
コラーゲンが生成されるメカニズムを見てみましょう。たとえば、血管の破裂を考えてみましょう。この破裂には、血管を閉じて血餅を形成することを目的とした無数の信号が伴います。
凝固には少なくとも30の特殊な酵素が必要です。血餅の後、組織の修復を続ける必要があります。傷に近い細胞もコラーゲンを生成します。これを行うには、最初に遺伝子の発現が誘導されます。つまり、遺伝子の情報から開始してタンパク質を生成できる生物です(遺伝子情報は、核を離れてリボソームに到達するmRNAに転写されます)遺伝子情報がタンパク質に変換される細胞質)次に、コラーゲンはリボソームで合成され(約1200のアミノ酸で構成され、分子量が約150,000 dの左側のらせんのように見えます)、その後、内腔に蓄積します。これは、翻訳後の修飾(「mRNAによって翻訳される言語の修飾)」を実行できる酵素の基質になります。コラーゲンでは、これらの修飾は、いくつかの側鎖、特にプロリンとリジンのヒドロキシル化で構成されます。
これらの変化を引き起こす酵素の失敗は壊血病を引き起こします:それは最初に血管の破壊、歯の破壊を引き起こし、その後腸間出血と死を引き起こす可能性がある病気です。それは長寿命の食品の継続的な使用によって引き起こされる可能性があります。
続いて、他の酵素の作用により、プロリンとリジンのヒドロキシル基のグリコシル化からなる他の修飾が起こります(糖はOHの酸素に結合します)。これらの酵素は内腔以外の領域に見られるため、タンパク質は修飾を受けながら小胞体内に移動し、小胞体に閉じこめられて小胞体から分離します。その中には、グリコシド化されたプロが含まれています。 -コラーゲンモノマー;後者は、特定の酵素がグリコシド化プロコラーゲンのカルボキシ末端部分に存在するシステインを認識し、異なる鎖を互いに接近させてジスルフィド架橋を形成させるゴルジ装置に到達します:このようにして、グリコシド化されたプロコラーゲンが互いに結合して得られ、これが出発点であり、相互に浸透し、次に自発的に三重らせんを生じます。一緒に結合されたグリコシド化されたプロコラーゲンの3つの鎖が到達し、次に小胞に到達し、それ自体を窒息させ、ゴルジ装置からそれ自体を切り離し、3本の鎖を細胞の周辺に向かって輸送します。原形質膜とイオン化すると、トリメーターは細胞から排出されます。
細胞外空間には、特定の酵素であるプロコラーゲンペプチダーゼがあり、細胞から排出された種から、300アミノ酸の3つのフラグメント(各ヘリックスに1つ)を1つ、末端のカルボキシ部分から、3つを除去します。アミノ末端部分からのそれぞれ約100アミノ酸のフラグメント(各ヘリックスに1つ):ヘリックスあたり約800アミノ酸からなる三重らせんが残っています。 トロポコラーゲン.
Tropocollagenはかなり堅い棒の外観を持っています;異なる三量体は共有結合と結合してより大きな構造を与えます: ミクロフィブリル。ミクロフィブリルでは、さまざまな三量体が千鳥状に配置されています。多くのミクロフィブリルはトロポコラーゲンの束を構成します。
骨のコラーゲン繊維の間には、カルシウムとマグネシウムの硫酸塩とリン酸塩が沈着する間質空間があります。これらの塩はすべての繊維も覆っています。これにより、骨が硬くなります。
腱では、間質腔は骨よりも結晶が豊富ではありませんが、トロポコラーゲンよりも小さなタンパク質が存在します。これにより、腱に弾力性が与えられます。
骨粗鬆症は、カルシウムとマグネシウムの不足によって引き起こされる病気であり、トロポコラーゲン繊維の間質領域に塩を固定することを不可能にします。

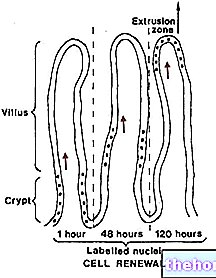
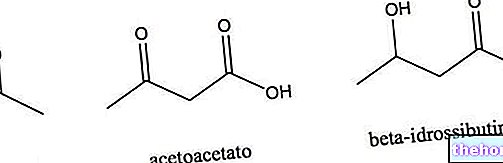
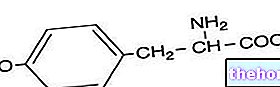
.jpg)











.jpg)




