遺伝的多様性がなければ、すべての生物は(遺伝によって)最初のものと同じでなければなりません。不平等な存在を持っているための唯一の説明は、単一の創造物に関連するものです。しかし、遺伝性の特徴の伝達の基礎となるDNAの構造は、絶対的な安定性ではなく、相対的な安定性を持っていることを私たちは知っています。安定性は初期情報の保存を保証しますが、不安定性は変更、またはより良い(特定の用語を使用するために)突然変異を決定します。
突然変異は3つの大きなグループに分けることができます:
-遺伝子変異;
-染色体変異;
-ゲノム変異。
この時点で、2つの概念を簡単に追加することをお勧めします。1つは「修復」の概念であり、もう1つは「好ましい」または「好ましくない」突然変異の概念です。
修復は、修復を意味する英語の用語であり、情報の保存を検証し、元のDNAに適合しないDNAの範囲を排除または修正することを目的とした、特定の酵素プロセスの細胞内の存在を指します。
「好ましい」または「好ましくない」突然変異の概念は、生物および種について、元の遺伝子(「野生」、すなわち「野生」、原始的)および突然変異遺伝子の効率の間の比較を表す。
長所と短所は常に特定の環境条件を指すことに注意してください。特定の環境での不利な突然変異は、さまざまな条件下で有利になる可能性があります。
この原則の結果は、正常またはマラリア環境でのヘモグロビン変異体の異なる生存効率によってすでに実証されているように、集団遺伝学において広大です。しかし、このトピックは、一般的、人間的、または医学的遺伝学に関するより詳細な記事に延期されるべきです。
体細胞変異と生殖細胞変異の間には、別の区別が必要です(特に遺伝子変異の場合)。私たちの多細胞生物のすべての細胞は、知られているように、単一の接合子に由来しますが、非常に早い段階で、全身が発達する体細胞系列と、生殖細胞系列を形成する運命にある生殖系列に分化します。配偶子2つの細胞集団間の相互作用を除いて、体細胞系列の突然変異は単一の生物に現れますが、子孫には影響を与えませんが、生殖細胞系列の突然変異は子孫でのみ発生します。
さまざまな種類の突然変異と相対的な結果に関して、突然変異の原因の分類について再度言及する価値があります。これらの原因は変異原と呼ばれ、主に物理的変異原と化学的変異原を区別するために使用されます。物理的環境のさまざまな変更は突然変異を引き起こす可能性がありますが、主な物理的突然変異誘発物質は放射線です。このため、放射性物質は危険であり、何よりも重い放射性粒子が亡命による突然変異を引き起こし、最も深刻な結果をもたらす傾向があります。
化学変異原は、核酸の秩序構造を変えることと、通常の窒素塩基と同様の物質を細胞に導入することの両方によって機能し、核酸の合成において後者と競合し、置換によって変異を引き起こす可能性があります。
遺伝子変異
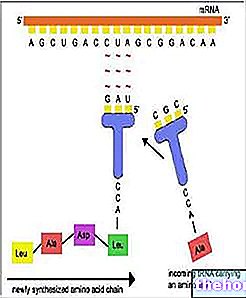
遺伝子変異は、1つまたはいくつかの遺伝子、つまり限られた範囲のDNAに影響を及ぼします。情報はヌクレオチドペアのシーケンスに保存されるため、最小の変異ユニット(ミュートン)は単一の相補的な塩基対で構成されます。遺伝子レベルでのさまざまな変異メカニズムについて詳しく説明しなくても、2つに限定できます。塩基置換塩基置換変異では、1つまたは複数のDNAヌクレオチドが他のヌクレオチドに置き換えられます。エラーが時間内に修正されない場合、RNAでも変更された転写時に配列が続きます。トリプレットは同義語に限定されず(遺伝子コードを参照)、ポリペプチド配列の1つまたは複数のアミノ酸が置換されます。アミノ酸の置換は、タンパク質の構造とその機能の決定にとって多かれ少なかれ重要である可能性があります。
再選択または挿入変異では、1つまたは複数のヌクレオチドが削除されるか、DNA配列に追加されます。これらの変異は、(単一のアミノ酸を加算または減算するトリプレット全体でない限り)読み取り順序で続くすべてのトリプレットが変更されるため、一般に非常に深刻です。
遺伝子変異は最も頻繁であり、個人間の遺伝形質の変動のほとんどの原因となっています。
染色体突然変異
これらは、染色体の比較的長い断片に影響を与える突然変異です。それらを主に次のように分類するのが通例です。
-再選のための染色体突然変異;
-重複による染色体突然変異;
-転座による染色体突然変異。
離反による突然変異は、多かれ少なかれ長い染色体断片の破壊と喪失によって起こります。特に減数分裂では、この種の突然変異は、多かれ少なかれ不可欠な特定の数の遺伝子が完全に失われるため、しばしば致命的です。
重複突然変異では、休憩後、染色分体の断端が融合する傾向があります。
その後のセントロメアの除去では、二動原体になった染色体がしばしば不均等な部分に分裂します。ご覧のとおり、結果は一方では再選され、他方では重複します。
染色体の切断の後に逆転が続く可能性があります。総遺伝物質は変更されていませんが、染色体上の遺伝子の配列が変更されています。
転座の場合も同様ですが、染色体の断片が非相同染色体に結合することに関係しています。一方の染色体は切断され、もう一方の染色体は長くなります。細胞の総遺伝情報は変わりませんが、位置効果はさらに顕著です。遺伝子作用の調節の概念を参照することで、位置効果を簡単に表すことができます。染色体上の位置にある遺伝子は、あるオペロンを簡単に離れて別のオペロンに挿入され、「活性化または抑制の変化」をもたらします。
しかし、2対の染色体間に転座の相反性があり、遺伝子情報の合計が変わらない場合、転座はバランスが取れている(またはバランスが取れている)と言われています。減数分裂のダイアキネシスの十字型の図は、一般的にバランスの取れた転座に対応します。
ゲノム変異
ゲノムは染色体で順序付けられた個々の遺伝的遺産であることを思い出して、染色体が種の標準とは異なる分布を持っている場合、ゲノム突然変異について話すことを指定できます。
ゲノム変異は、主に倍数性、半数性、異数性の変異に分類できます。
倍数性突然変異は、重複が分裂を引き起こさない場合に発生します。それらは植物でより簡単に発生し、実際にそれらは生産を改善するために使用されます。
細胞分裂が有糸分裂に失敗した場合、二倍体配偶子が発生する可能性があります。そのような配偶子が正常な配偶子と融合することに成功した場合、この受精から生じる接合子は三倍体になります。複製と有糸分裂は偶数の染色体を必要としないので、そのような接合子は時々生物全体を生み出すことに成功することができます。しかし、減数分裂の時点では、相同染色体の定期的なペアリングは不可能です。
一倍体によるゲノム変異は、通常は二倍体の種で、核物質を欠く別の配偶子によって配偶子が活性化される場合、または受精がない場合でも発生する可能性があります。一倍体個体が発生します。
以前のタイプのゲノム突然変異は常に染色体の整数nの加算または減算に関係しますが、異数性の突然変異は単一染色体の過剰または欠陥(染色体異常)に関係します。
種の正倍数体の染色体構成は、その正常な核型またはイディオタイプとして定義されます。