体温調節は生物学的メカニズムの統合システムであり、生物の外部の気候条件に関係なくほぼ一定の内部温度を維持する役割を果たします。これらのメカニズムは、鳥や哺乳類(すべての恒温動物)では特に効果的ですが、魚、両生類、爬虫類ではそれほど効果的ではありません(ポイキロサーミックアニマル)-のプロセスを含む 製造, ストレージ と 分散 熱の。
肥満の被験者は、他の正常体重の人と比較して異常な食事をしないことが多いため、同じ身体活動で、体温調節プロセスの変化がエネルギー消費の削減につながる可能性があります。脂肪の形での過剰なエネルギーの蓄積したがって、肥満とは異なり、痩せた被験者は、熱の形で過剰な食物(茶色の脂肪組織を参照)を処分するのに優れています。
体温調節は、何よりもまず自発的または非自発的である可能性があります。最初のケースでは、要素から保護された巣穴の探索や、自分の体温を維持するのに最も適した場所への移動など、適切な行動戦略を自発的に開始するのは動物自身です。

不随意の体温調節反応でさえ、寒い環境や暑い環境にさらされることで引き起こされる可能性があります。いずれの場合も、視床下部の体温調節センターの介入が必要であり、皮膚および中枢の体温受容体(脳、脊髄にある)からの信号をキャプチャして処理することができます。脊髄および中枢器官)、体温を維持するのに最も適した生理学的反応を調整します。
寒冷環境での体温調節
体温調節の寒さへの適応は、熱を保存および/または生成することを目的としています。
生物が熱を発生する能力は、熱発生と呼ばれます。それは主に必須であり、食事とともに導入された栄養素の移動、消化、吸収、および処理に関与する生理学的および代謝プロセスに関連しています。
哺乳類は、スリルメカニズムに関係するかどうかに関係なく、熱産生を増加させる能力があります(オプションの熱発生)。最初のケースでは、震える熱発生について話します。このメカニズムは、運動を目的としたものではなく、筋肉組織のリズミカルで等尺性の収縮を介して熱を発生させます。収縮と弛緩が交互に起こると、震えと呼ばれる特徴的な震えが起こります。これは、体温が「著しく」低下する傾向があるときに現れます。震えは、安静時の筋肉よりも最大6〜8倍の熱を発生します。 、最大の血管収縮(下記参照)が体温を維持できなかった場合にのみ発生します。
化学的熱発生とも呼ばれる非スリル熱発生には、発熱(熱を発生する)生化学反応による熱の生成が含まれます。これらの反応は、褐色脂肪組織(BAT)、肝臓、筋肉などの特定の臓器で発生します。
したがって、冬眠中の動物に典型的で、人間にはほとんどない(乳児では多い)褐色脂肪組織は、ミトコンドリアレベルに存在するカロテノイドによって与えられる特徴的な褐色色素沈着(肉眼で見える)に対して定義されます。褐色のこれらのエネルギー中心脂肪細胞は、ミトコンドリアタンパク質UCP1の存在というさらなる特徴で区別されます。ミトコンドリア膜のレベルに位置するこのタンパク質は、酸化的リン酸化を分離するという特徴を持っているため、 ATP分子、褐色脂肪組織は、熱の生成を増加させるために栄養素(主に脂肪)を燃焼させる目的を持っています。寒さによって刺激された褐色脂肪組織の活性化は、主にノルアドレナリンの放出とその相互作用に関連していますβ3受容体を伴うが、T3eの放出などの内分泌メカニズムによっても保証される甲状腺からのT4。褐色脂肪組織の最大の沈着物は、肩甲骨間、大動脈周囲および腎周囲の領域に記録されています。これらのレベルでは、それらは血管の近くにあり、そこに熱を放出して、血流とともに体の周辺領域に輸送されます。
現在、肝臓は体温調節にも関与しており、人体が低温にさらされると代謝活性が高まり、熱が発生すると考えられています。もう1つの最近の発見は、筋肉内のUCP1タンパク質のアイソフォームの発見でした。これは、代謝起源のサーモゲニンの役割の疑いを示唆しています(震えによって熱を生成する能力に加えて)。これらの状況(BATなど)での活動組織の代謝要求をサポートし、すべての解剖学的領域でそこで生成される熱の輸送を増加させるために必要です。これらすべてを保証することに加えて、心臓活動の増加はそれ自体で可能です。無視できない量の熱を生成します。
熱損失の制御は、伝導、対流、放射、および蒸発の物理法則によって管理されます。
伝導:表面を介して互いに接触している、異なる温度での2つのオブジェクト間の熱伝達。
輻射または輻射:接触していない異なる温度での2つのオブジェクト間の熱伝達。熱の損失または獲得は、可視または赤外線範囲の波長の放射の形で発生します。明らかに、太陽が宇宙を介して地球を加熱するのと同じ方法です。によって失われる熱量の半分以上を構成します。人体。
対流:物体からその中を移動する熱源(空気または水の流れ)への熱の伝達。暖かい肌を通る水または冷たい空気の動きは、熱の継続的な除去を引き起こします。
蒸発:発汗によって失われた液体の液体から気体状態への通過による熱伝達、皮膚および気道を介した鈍感な損失。
環境中の熱分散の減少は、本質的に皮膚の血流の封じ込め(血管収縮)と立毛(毛皮の動物では、暖かい皮膚と冷たい環境の間に、断熱材として機能するエアクッションが作成されます)によって発生します。
食欲の増加は、その一部として、食事によって誘発される熱発生メカニズムを介して熱の生成を増加させ、熱発生器官のエネルギー需要をサポートします。
暑い環境での体温調節
暖かい環境にいる間、生物は一連の熱分散メカニズムを介して反応しますが、多くの点で今説明したものとは逆です。さらに、オプションの熱発生の根底にある代謝プロセスの中断があります。これらの中で、皮膚の血管拡張と増加を覚えています。発汗、呼吸の頻度と深さ(ポリプネア)の、蒸発による熱の分散を高めることを目的としたすべてのプロセス。これらの状況では、熱発生器官による酸素の需要の低下に応じて、食欲と心拍数も低下します。
長期的な適応プロセスの中で、甲状腺刺激ホルモンの下垂体分泌の減少を認めることも可能であり、その結果、代謝が遅くなり、したがって熱が発生します。
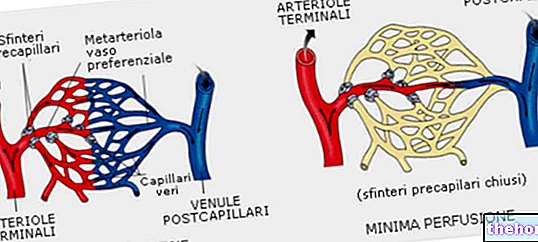
前の章で述べたように、血管収縮プロセスは主に交感神経系によって制御されています。前毛細血管括約筋および細動脈の平滑筋は、節後交感神経(アドレナリン作動性)ニューロンから入力を受け取ります。深い温度が下がると(寒さにさらされると)、視床下部はこれらのニューロンを選択的に活性化し、ノルアドレナリンの放出によって細動脈平滑筋の収縮を決定し、皮膚の血流を減らします。この温度調節反応は、内臓への血液を暖かく保ちます。 。、天候によって冷えた皮膚表面の血流を最小限に抑える血管収縮は能動的なプロセスですが、血管拡張は主に受動的なプロセスであり、交感神経活動を阻害することによって血管収縮剤の活動を停止することに依存します。このプロセスが交感神経の典型的な場合活動。体の四肢、体の他の部分では、血管拡張は、アセチルコリンを分泌する特殊なニューロンによって支持されます。特別な場合は、一酸化窒素(NO)または他の血管拡張パラクリン物質の放出後のいくつかの血管領域の局所拡張によっても表されます。
体温調節の文脈では、皮膚の血流は、熱を節約する必要がある場合のゼロに近い値から、熱を環境に放出する必要がある場合の心拍出量のほぼ1/3まで変化します。