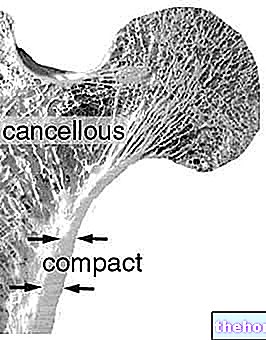
ステファノカザーリ博士が編集
血管の収縮または拡張の状態の制御は、その厚さが平滑筋を有する構造でのみ実行することができます。この制御は、神経、ホルモン、代謝に由来する可能性があるため、遠隔または局所制御です。灌流された臓器の重要性または生理学的特性に応じて、すべての循環器系は、ある作用機序が別の作用機序よりも優勢になります。筋肉のチュニックがない毛細血管では、壁の状態は毛細血管前括約筋に厳密に依存していますが、とりわけ経壁圧に依存しています。
血管神経制御
神経起源の血管収縮は、化学伝達物質(ノルアドレナリン)を介して筋肉に作用し、血管収縮を誘発する交感神経アドレナリン作動性血管収縮剤の作用に依存します。交感神経血管収縮系の作用は一定であるため、血管緊張、しかし特に動脈では、それは拡張圧を決定する責任があり、末梢抵抗に直接作用します。静脈還流の封じ込め血管、静脈および洞を通して。頭のシステムに作用するようには見えません。
- 線維はT1-L4管の中間側柱から始まり、白い連絡枝の形で出て、交感神経節鎖の構成に入り、求心性神経の構成に入ります。
血管拡張は、交感神経アドレナリン作動性の抑制に依存する受動的であるか、または直接的または間接的作用で能動的である可能性があります。
- 住む:感覚神経の刺激は、コリン作動性作用による外分泌腺の血管拡張の場合のように、キニンの産生を誘発します。また、顎下腺への作用を伴う鼓膜の索神経の刺激の特定の場合のように、カリクレイン-カリジン-ブラジキニンの産生。
- 直接:アセチルコリン、ドーパミン、ヒスタミンなどのメディエーターの作用に基づく。血管の筋肉組織に。それは交感神経または副交感神経の起源である可能性があり、自律神経系線維の起源であるS2-S4管の除去が勃起を損なうのではなく、反射のみを損なう、亀頭神経の場合のように2つのシステムが統合されることがよくあります亀頭の刺激に由来するもの。
直接的な血管拡張作用は、バロおよび化学受容体の刺激による圧力の反射調節に介入せず、頭側領域では決定的な作用はありません。特徴的ですが、完全に仮説的な作用は、骨格筋のコリン作動性交感神経系に起因します。視床下部刺激に続いて、高ストレスの状態で認められるびまん性血管拡張によって強調されます。
軸反射:これは、感覚神経の末梢断端の刺激に続く、Cニューロンによって媒介される反射型応答であり、したがって、血管拡張を引き起こす脊髄中心を伴わない。したがって、衝動は中央で進行して痛みの情報を伝達し、遠心力で血管拡張を誘発します。このメカニズムは、皮膚の三重反応の根底にあります。
カテコールアミン
ノルアドレナリン:交感神経のメディエーターおよび動脈内注入の両方として、血管収縮剤としてのみ機能します。
アドレナリン:脾臓、腎臓、皮膚の血管収縮剤、冠循環の血管拡張剤、肝臓、骨格筋です。大量のアドレナリンは、アルファ受容体とも相互作用するため、一般的な血管収縮を引き起こします。いずれにせよ、循環カテコールアミンの効果は、交感神経によって媒介される効果よりも明らかに少ないです。
アルファ受容体:それらはノルエピネフリンとのみ相互作用し、正の変力作用を有する心臓にはほとんど存在しません。血管平滑筋に大量に存在します。
Beta1受容体: それらは心臓の両方のカテコールアミンと相互作用し、上記の受容体などのカルシウムイオンの移動度を増加させることにより、変時作用、変伝導作用、および陽性変力作用を誘発します。
Beta2受容体:肝臓、心臓、骨格筋に存在し、腎臓、脾臓、皮膚には存在しません。
アンジオテンシン: レニンの作用によりアンギオテンシノーゲンの誘導体である全身性低血圧で合成され、抵抗性血管にのみ作用し、持続時間が短い。
バソプレッシン: 視床下部の視索上核によって産生され、全身の抗利尿作用と血管収縮作用があり、毛細血管前括約筋、抵抗血管だけでなく細静脈にも作用します。
オータコイド
ヒスタミン: 肥満細胞に含まれるものは、外傷後に放出され、細動脈の血管拡張、局所静脈領域の血管収縮を引き起こし、毛細血管透過性を増加させます。骨格筋では、オルソシンパシートーンの低下によりそれらも放出されます。
セロトニン: 血小板の凝集がないため、損傷した血管の血管収縮を引き起こします。胃の中では、それらの分泌はガストリンによって引き起こされます。それらはアドレナリン受容体を遮断し、細動脈の血管拡張と細静脈の収縮を引き起こして間質液の利用可能性を高めます。
血管拡張性代謝物:
全身性充血は、単一のイオンまたは代謝物に起因することはできませんが、灌流される組織の生理機能に常に従う全体に起因しますイオンカリウム、カルシウム、しかしとりわけ酸素分圧の変動、または高炭酸ガス血症、増加を伴わないしかし、血流では、代謝物によって誘発される血管拡張の最も頻繁な原因です。明らかに、これらのシステムには局所的な作用があります。神経質:実際、正副交感神経系は心臓の緊張の基部にあり、循環系にはあります。オルソシンパシー起源の収縮トーンのみ。拡張は、反射に基づいて血管運動の抑制に起因します。アドレナリンの「作用」によって強制できるのは一部の領域のみです。
統合された体圧調節システム:
数秒:
- バロセプティブシステム
- CNSの虚血メカニズム
- 化学受容器のメカニズム
秒から分:
- レニン-アンジオテンシン系
- 応力緩和メカニズム
- キャピラリーを通る液体の動きのメカニズム
無限大までの分:
- レニン-アンギオテンシン-アルドステロン系によって統合された腎液系
参考文献:
- Stagnaro-Neri M.、Stagnaro S.、 生物物理学的記号論:動脈コンプライアンスと末梢動脈抵抗の評価。 XVIIコンの行為。ナット。 Soc。Ital。MicrocirculationStudio、フィレンツェ1995年10月、BibliotecaScient。ミリタリーヘルススクール、2、93。
- ファイファーJR。 下肢の静脈系の解剖学と生理学。 Ed Phlebologle、1992年。
- Braundawall E.心臓病: 循環器内科の論文。エド・ピクシン。
- 林K .. 動脈壁の機械的特性と構成則を測定するための実験的アプローチ、Journal of Biomechanical Engineering、Vol.115。
- TestutL.。人体解剖学、4冊目の本:脈管学。
- ATM。アトラステキスト- 基本的な概念。ランペッロA。
- タグリエッティ-ボックス 「生理学の原則」、GoliardicaPavese。
- シルバーソーン "生理"、アンブロジアーナ出版社。
- De Trafford J. C.、Lafferty K.、Kitney R. I.、Cotton L. T.、Roberts V.C.人間の血管運動制御システムのモデリングと動脈疾患の調査へのその応用。 IEEEProc。129A、1982。
- グリーンJ.H.人類生理学入門。 Zanichelli、1972年。
- ガイトンA.C.医学生理学に関する論文。教授によるVアメリカ版のIIイタリア版。アルフレド・クラトロ、ピクシン・ヌオーヴァ・リブラリア、パドヴァ、1987年。
- Montano N.、Gnecchi Ruscone T.、Porta A.、Lombardi F.、Pagani M.、MallianiA。段階的な起立性傾斜中の交感神経バランスの変化を評価するための心臓変動のパワースペクトル分析。循環、vol。 90、n°4、1994。
- バートンA.C.循環の生理学と生物物理学。紹介文。イタリア語版博士によるフランコ・トリポーディ、科学思想出版社、ローマ、1983年。