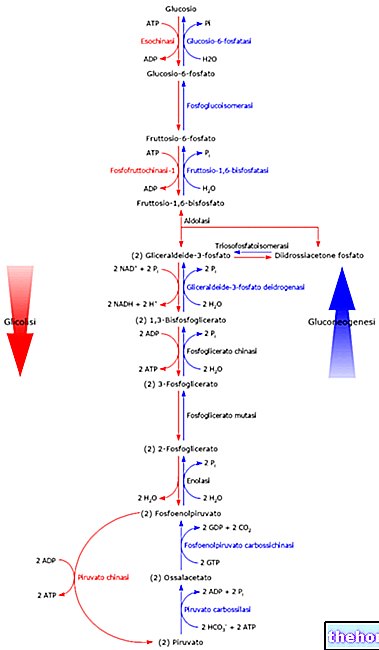
細胞が嫌気性条件で機能する場合、グルコースを乳酸に変換することによってエネルギーを生成し、コリ回路を介して後者を処分します。利用可能な酸素がある場合(したがって休止状態で)、グルコースの90%以上が好気的に消費されます。嫌気的に残りの10%だけ。好気性経路が提供できるよりも多くのATPが必要な場合(たとえば、筋肉にストレスがかかっている場合)、嫌気性代謝によって追加の供給が提供されます(酸素不足の状態にあります:息切れ、疲労呼吸などで):糖新生によって乳酸塩(解糖から得られる)をブドウ糖に変換することにより、この代謝を加速する必要があります。
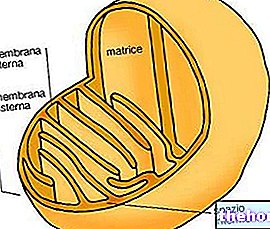
ミトコンドリアでは有酸素代謝が起こります。
有酸素代謝で最初に遭遇する酵素は ピルビン酸デヒドロゲナーゼ;ピルビン酸デヒドロゲナーゼは、3つの触媒部位が連続して作用する48〜60のタンパク質単位の集合体であるため、酵素ではなく酵素複合体であると言った方が正確です。
ピルビン酸デヒドロゲナーゼは、次の反応(レドックス)を触媒します。
ピルビン酸+ NAD + + CoA-SH→アセチルCoA + NADH + H + + CO2
CoA-SHは補酵素Aです:それはパントテン酸の誘導体です;アセチル補酵素Aはチオエステルです。ピルビン酸の最初の炭素が酸化数3から酸化数4(酸化)になり、ピルビン酸の2番目の炭素が酸化数2から酸化数3(酸化)になるため、これはレドックスプロセスです。次に、ピルビン酸が酸化され(2つの電子が完全に失われます)、NADが還元されます。
前述のように、ピルビン酸デヒドロゲナーゼには3種類の酵素活性があり、それぞれが独自の触媒補因子によってサポートされています。
- チアミンピロリン酸(ビタミンB1の誘導体です);それは脱プロトン化された形で活性です:カルバニオンが形成されます。
- リポアミド(リポ酸の誘導体);それは非常に反応性の高いジスルフィド架橋を含んでいます。
- フラビンアデニンジヌクレオチド(ビタミンB2の誘導体です);それは酸化還元特性を持つヌクレオチドです:その酸化還元中心はフラビンで構成されています。
真核細胞では、好気性代謝はミトコンドリアである細胞の特殊な細胞小器官で起こります。バクテリアでは、ブドウ糖や他の種の代謝が細胞内で起こりますが、特殊な細胞小器官はありません。
ピルビン酸がミトコンドリアに入ると、「糖新生を実行する必要がある場合はピルビン酸カルボキシラーゼの作用(出発物質を再構築するため)、またはエネルギーを生成する必要がある場合はピルビン酸デヒドロゲナーゼを行うことができます。 「好気性代謝によって形成されるアセチル補酵素Aは、ピルビン酸カルボキシラーゼの作用を刺激するため、糖新生を促進し、ピルビン酸デヒドロゲナーゼの作用を低下させます。
ピルビン酸デヒドロゲナーゼがどのように機能するかを見てみましょう。まず第一に、チアミンピロリン酸の作用によるピルビン酸の脱炭酸があります。
酸性環境では、陰イオン型のチアミンピロリン酸が活性であり、酸性pHでプロトン化され、脱炭酸が起こらないため、好気性代謝を阻害する可能性があります。
炭素-炭素結合を切断する必要があるため、脱炭酸は難しい反応です。この場合、反応中間体(ヒドロキシエチル-チアミンピロリン酸)が共鳴を与える(分子のp電子が非局在化する)という事実によって、反応は熱力学的に有利になります:ヒドロキシエチル-チアミンピロリン酸は(共鳴の)3つの可能な形態で存在しますこれにより、非常に安定します。さらに、陰イオン形態のヒドロキシエチル-チアミンピロリン酸は、リポアミドのジスルフィド架橋(ピルビン酸デヒドロゲナーゼの第2の触媒補因子)と相互作用することができるのに十分長い時間生存します;ジスルフィド架橋は振動アームです(それは長い柔軟な鎖の末端)であり、酵素複合体のある触媒部位から別の触媒部位に移動することができます。
次に、リポアミドは、ジスルフィド架橋を介して、ヒドロキシエチル-チアミンピロリン酸に結合します。アセチルリポアミドが得られます。今説明したものは、ピルビン酸デヒドロゲナーゼ複合体の最初の酵素によって触媒されるトランスアセチル化反応の最初のステップです。この段階で、ヒドロキシル基とピロリン酸チアミンとの間の結合が切断され、元の形態に戻りました。酸化還元反応が起こり、ジスルフィド架橋が酸化剤として作用し(2つの硫黄原子が還元され)、ヒドロキシル基の比較でアセチルに酸化された。
この段階の後、リポアミドの振動アームが動き、ピルビン酸デヒドロゲナーゼの第2酵素に近づき、アセチル基を運ぶことによって真のトランスアセチラーゼ活性を実行します。第2酵素によって触媒されるトランスアセチル化反応の第2段階が起こります。このようにして、アセチル補酵素Aを取得しました。還元型のリポアミドを復元する必要があります。ピルビン酸デヒドロゲナーゼの3番目の酵素が介入し、リポアミドを酸化還元して、その電極をFADに転送します。FADはFADH2に還元されます。 FAD / FADH2は、2つの異なるモノエレクトロニックステージまたは単一のバイエレクトロニックステージでレドックスペアとして機能できます。
FADH2はすぐにその電子をNAD +に与え、FADとNADH + H +を取得します。
記載されているように得られたアセチル補酵素Aは、クレブス回路(またはトリカルボン酸回路)の出発生成物です。