有糸分裂中の細胞が、有糸分裂または抗有糸分裂毒と呼ばれるコルヒチンなどの物質の作用、またはスタトモキネティクスにさらされると、紡錘体のセントロメアの移動メカニズムがブロックされ、染色体は中期段階にとどまります。 、写真を撮り、染色体を拡大し、明確に定義された分類基準(セントロメアの相対位置とサイズ)に従って順番に並べます。こうして各細胞の核型が得られます。平均値(単一のエラーを避けるため)は次のようになります。核型個体。
最も簡単に検出できる染色体異常は、トリソミー、一染色体、またはヌルアイソミーによるものです。これらはそれぞれ、特定のタイプの染色体が3つ、1つ、またはまったく存在しないことに対応します。 1つまたは3つの染色体の存在は、しばしば非常に深刻な形態学的および代謝的変化とはいえ、独立した生活と両立することができ、投与量と遺伝子間の相互作用の通常のバランスを変えます。 nullisomyは、染色体ペアに含まれるすべての遺伝子の完全な欠如を補うことができないため、致命的と見なされます。
トリソミーの典型的なケースは、一般に「モンゴリズム」として知られている「トリソミー2」またはダウン症です。
一染色体性の症例は、これから見ていくように、ゴノソームが1つしかないターナー症候群(X)で発生します。
転座による異常と減数分裂不分離による異常を区別することが可能であり、原因、影響、遺伝的伝達、そしてその結果としての優生学的予後は、細胞学的緩和と同様に異なります。
細胞質の遺産
これまでに行われた遺伝学のすべての治療は、核DNAの挙動、その伝達、細胞質および外部環境との関係の研究に基づいています。
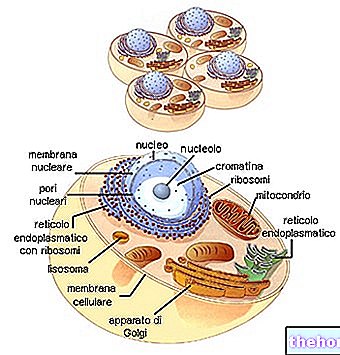
遺伝における核の役割は絶対に最重要ですが、細胞質にさまざまな方法で関連する遺伝的伝達の現象もあります。たとえば、独自の遺伝的連続性を持つ細胞質オルガネラ(ミトコンドリア、色素体、中心)は、別個の遺伝的伝達現象を示す可能性があります複雑な相互作用はあるものの、核からのもの。母体が初期発生のために胚に供給する予備物質の含有量でさえ、細胞質遺伝の現象を決定することができます。一般に、遺伝性の遺伝の発現はさまざまであると言えます。細胞質および細胞外環境の要因との相互作用によって条件付けられます。