ジョバンニ・チェッタ博士が編集
生化学から生体力学へ

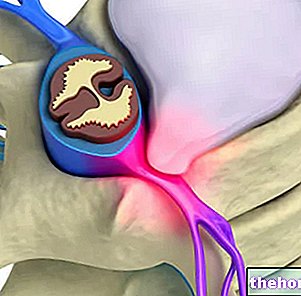
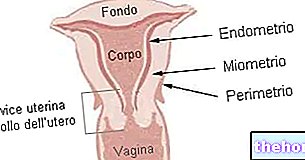
細胞外マトリックス(MEC)
今日私たちがほとんど知らないことではありますが、ECM(細胞外マトリックス)の説明は、健康における脊椎と姿勢の変化の重要性をよりよく理解するために不可欠です。
すべての細胞は、すべての多細胞生物と同様に、その重要な機能を実行して生き残るために、その環境を「感じ」、相互作用する必要があります。多細胞生物では、細胞は人間のコミュニティのようにさまざまな行動を調整する必要があります。

ECMは一般に、いくつかの大きなクラスの生体分子で構成されていると説明されています。
- 構造タンパク質(コラーゲンとエラスチン)
- 特殊なタンパク質(フィブリリン、フィブロネクチン、ラミニンなど)
- プロテオグリカン(アグリカン、シンデカン)およびグルサミノグリカン(ヒアルロン酸、コンドロイチン硫酸、ヘパラン硫酸など)
構造タンパク質の中で、私は コラーゲン 動物界で最も代表的な糖タンパク質のファミリーを形成します。それらはECMで最も存在するタンパク質であり(最も重要ではありません)、適切な結合組織(軟骨、骨、筋膜、腱、靭帯)の基本的な構成要素です。
コラーゲンは主に線維芽細胞によって合成されますが、上皮細胞もそれらを合成することができます。
コラーゲン繊維は、細胞の寿命の基本となる生物学的連続体を構成するECMの他の膨大な量の分子と継続的に相互作用します。原線維内の関連するコラーゲンは、張力に抵抗できる構造の形成と維持において主要な役割を果たします。非弾性コラーゲンは、機械的負荷の関数として生成および再代謝され、その粘弾性特性は人間の姿勢に大きな影響を与えます。
PG / GAGコーティング(プロテオグリカン/グルコサミノグリカン)のおかげでコラーゲン繊維は、バイオセンサーと生体伝導特性を備えています。実際、構造変形を発生させることができる機械的な力は、分子間結合に応力を加えてわずかな電束、つまり圧電電流を生成することを私たちは知っています(Athenstaedt、1969)。したがって、三次元で遍在するコラーゲンネットワークは、コラーゲン原線維と細胞の間の相対的な配置に基づいて、求心性方向(ECMから細胞へ)またはその逆で、空間の三次元で生体電気信号を伝導するという独特の特性も持っています。逆に遠心性。
これはすべてリアルタイムのMEC細胞通信システムを表しており、そのような電磁生体信号は重要な生化学的変化を引き起こす可能性があります。
最後に、細胞は、当然のことながら、継続的かつかなりのエネルギー消費(約70%)の物質を生成し、特定の小胞(コラーゲンの生物学的前駆体)の排他的貯蔵を通じて必然的に排出されなければならないことを強調する必要があります(アルベルガティ、2004)。
脊椎動物の組織の大部分は、強度と弾力性という2つの重要な特性が同時に存在する必要があります。の実際のネットワーク 弾性繊維これらの組織のECM内にある、は、強い牽引力の後に初期状態に戻ることができます。弾性繊維は、臓器またはその一部の伸展性を少なくとも5倍に高めることができます。長くて弾力性のないコラーゲン繊維が弾力性のある繊維の間に散在し、「組織の牽引による過度の変形」を制限するという正確な役割を果たします。エラスチンは弾力性のある繊維の主成分であり、弾力性のある血管に特に豊富に見られます(靭帯、肺、皮膚の大動脈の総乾燥重量の50%以上を占めます。平滑筋細胞と線維芽細胞は、その前駆体であるトロポエラスチンの主要な産生物質です。
ECMには、他のECM分子や細胞表面受容体に特異的な結合部位を持つ特殊な非コラーゲンタンパク質が多数含まれています(まだ十分に定義されていません)。このように、これらのタンパク質のすべてのコンポーネントは、類似分子と異なる分子の両方の間の接触の「増幅器」として機能し、数百万および数百万の生化学的情報を生成、変調、変化、および伝播できる無限の生化学的ネットワークを作成します(そしてエネルギー)。
重要な「細胞外マトリックスの特殊なタンパク質は フィブロネクチン、すべての脊椎動物に見られる高分子量糖タンパク質。フィブロネクチンは、細胞増殖、細胞間接着、およびECMにより、さまざまな方法で細胞移動に影響を与えることができるようです(細胞は1日あたり最大5 cm移動できます-Albergati、2004)。最もよく知られているアイソフォームであるタイプIIIはインテグリンに結合します。後者は、機械受容体として機能する膜貫通タンパク質のファミリーです。これらは、選択的かつ変調可能な方法で、細胞内のECMから機械的牽引と押し出しを伝達し、その逆も同様であり、細胞骨格に関与する一連の反応を細胞質に誘導します。それらが細胞接着、成長および移動を調節する他のタンパク質(Hynes、2002年)。
グリコサミノグリカン(GAGS)とプロテオグリカン(PG)は、結合組織内に定義された高度に水和したゲル状の物質を形成し、その中にフィブリルタンパク質が収容され、覆い隠されています。この形態の多糖類ゲルは、一方ではECMがかなりの圧縮力に抵抗することを可能にし、他方では血液と組織の間の栄養素、代謝物およびホルモンの迅速で一定の選択的な拡散を可能にすることができます。
グルコサミノグリカンの多糖鎖は、体積的に硬すぎて、ポリペプチド鎖に典型的なコンパクトな球状構造内で折りたたむことができず、さらに親水性が高いため、GAGは極端なコンフォメーションをとる傾向があります。質量に比べて大きな体積を占めるため、低濃度でもかなりの量のゲルを形成します。大量の負電荷(GAGは最も多くの陰イオン細胞を表し、通常は硫酸化され、動物細胞によって生成されます)は多数の陽イオンを引き付けます。これらの中で支配的な役割はNa +によって果たされ、それは全体の浸透能力を与え、ECMに膨大な量の水を閉じ込めます。このようにして、ECMが重要な圧縮力にさえ対抗できるようにする腫れ(膨圧)が生成されます(これのおかげで、たとえば、股関節の軟骨は、生理学的条件下で、数百気圧の圧力に完全に耐えることができます)。
結合組織内では、GAGは全重量の10〜12%未満ですが、その特性により、さまざまなサイズと密度の電荷の水和ゲルの細孔を形成する細胞外空間の多くを満たし、選択的に機能します。 MEC内の分子と細胞のトラフィックが、それらのサイズ、重量、および電荷に基づいて規制されるキーポイントまたは「サーバー」。
ヒアルロン酸(ヒアルロン酸、ヒアルロン酸)は、おそらく最も単純なGAGであり、実験的および分子生物学的データにより、かなりの圧力への耐性に関して、骨や関節のレベルで基本的な役割を果たしていることが確認されています。 :それは、後の段階でそれらが移動する細胞の間に空のスペースを作成します(Albergati、2004)。
すべてのPGがECMによって分泌されるわけではなく、一部は原形質膜の不可欠な構成要素です(Alberts、2002)。
したがって、細胞外マトリックスは、タンパク質、PGS、GAGが、あらゆる組織や有機活動の構造的サポートや調節など、無数の機能を提供する非常に複雑なネットワークと見なすことができます。グローバルな細胞の恒常性は、細胞の内部またはECMの外部で発生および発生するメカニズムの複合体と見なす必要があります。後者の場合、細胞は中間または最終のターゲットを表すことができます。細胞外成分は、細胞足場の物理的支持構造を表すことに加えて、細胞内環境と臓器およびシステムの両方に関する重要なプロセスの開始、発達、および終了のための実際の部位としても機能します。私たちは、数百万、数百万の情報を遠く離れた場所でも生成、変調、変化、伝播できる無限の生化学的ネットワークに直面しています。
体の各細胞は、機械的および化学的およびエネルギー的側面の両方で常にECMと相互作用し、組織の静的および動的アーキテクチャに「劇的な」影響を及ぼします。 P. A. Bacciによれば、間質マトリックスは、生命反応の母、まず第一に、物質とエネルギーの間の交換が行われる場所を真に表しています。すべての組織は、閉じたシステムではなく開いたシステムで相互に接続され、機能的に統合されています。それらの間で継続的な交換が行われ、それは、生化学的、生物物理学的、および電磁的メッセージを利用して、つまり、さまざまな形態のエネルギーを使用して、局所的および全身的に行うことができます。
F.Gとして収容されている細胞と細胞外マトリックスは、明らかに別々の2つの世界を表しており、正しく相乗的に機能するためには、生命の全期間にわたっていつでも相互作用する必要があります。これには、並外れた一連の信号と、それに続く同様に信じられないほどの一連の分子生物学的活動が必要です。
「細胞外マトリックス-構造と機能」に関する他の記事
- 脊柱側弯症の治療
- 脊柱側弯症-原因と結果
- 脊柱側弯症の診断
- 脊柱側弯症の予後
- 結合組織と結合筋膜
- コネクティブバンド-特徴と機能
- 姿勢とテンセグリティ
- 人間の動きと骨盤位サポートの重要性
- 正しい骨盤位と咬合サポートの重要性
- 特発性脊柱側弯症-払拭する神話
- 脊柱側弯症と治療プロトコルの臨床例
- 治療結果臨床例脊柱側弯症
- 自然な態度としての脊柱側弯症-参考文献