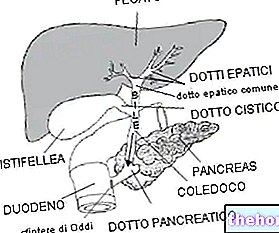
「」のわかりやすい方法で話すことができるようにするためにヘモグロビン(Hb)、最初の世話をするのは便利です ミオグロビン(Mb) これはヘモグロビンに非常に似ていますが、はるかに単純です。ヘモグロビンとミオグロビンの間には密接な親族関係があります。どちらも複合タンパク質であり、それらの補欠分子族(非タンパク質部分)がグループです。 ヘム.
ミオグロビンは、約150個のアミノ酸(生物によって異なります)の一本鎖からなる球状タンパク質であり、その分子量は約18Kdです。
前述のように、繊維状タンパク質のα-ヘリックス構造に起因する折り畳みからなる、タンパク質の疎水性(または親油性)部分に挿入されるヘム基を備えています。
ミオグロビンは主にα-ヘリックスのセグメントで構成され、8つ存在し、ほとんどが非極性残基(ロイシン、バリン、メチオニン、フェニルアラニン)で構成されていますが、極性残基は実質的に存在しません(アスパラギン酸、グルタミン酸、リジン)。およびアルギニン);唯一の極性残基は2つのヒスチジンであり、ヘムグループへの酸素の結合に基本的な役割を果たします。
ヘム基は発色団(可視光に吸収)であり、ミオグロビンの官能基です。
参照:糖化ヘモグロビン-尿中のヘモグロビン
少しの化学

プロトポルフィリンと鉄の間の結合は、中心原子(またはイオン)がその酸化数(電荷)よりも多い数で他の化学種と結合を形成する化合物である配位化合物の典型的な結合です。ヘムの場合、これらの結合は可逆的で弱いものです。
鉄の配位数(配位結合の数)は6です。鉄の周りに6つの分子が結合電子を共有している可能性があります。
配位化合物を形成するには、正しい向きの2つの軌道が必要です。1つは電子を「獲得」でき、もう1つは電子を供与できます。
ヘムでは、鉄はプロトポルフィリン環の中心にある4つの窒素原子と4つの平面結合を形成し、近位のヒスチジン窒素と5番目の結合を形成します。鉄は6番目の自由配位結合を持ち、酸素に結合できます。
鉄が遊離イオンの形であるとき、そのタイプの軌道 NS それらはすべて同じエネルギーを持っています。ミオグロビンでは、鉄イオンはプロトポルフィリンとヒスチジンに結合しています。これらの種は軌道を磁気的に乱します NS いくつかの鉄;摂動の程度は、さまざまな軌道で異なります NS それらの空間的配向と摂動種のそれによって異なります。軌道の総エネルギーは一定でなければならないので、摂動はさまざまな軌道の間にエネルギーの分離を引き起こします。いくつかの軌道によって獲得されたエネルギーは、他の軌道によって失われたエネルギーと同等です。
軌道間で発生する分離がそれほど大きくない場合は、高スピンの電子配置が望ましいです。結合電子は、可能な限り多くのサブレベル(最大多重度)で平行スピンに配置しようとします。一方、摂動が非常に強く、軌道間に大きな分離がある場合は、低エネルギー(低スピン)軌道で結合電子をペアリングする方が便利な場合があります。
鉄が酸素に結合すると、分子は低いスピン配列を取りますが、鉄が6番目の配位結合を持たない場合、分子は高いスピン配列を持ちます。
このスピンの違いのおかげで、ミオグロビンのスペクトル分析を通して、酸素(MbO2)がそれに結合しているかどうか(Mb)を理解することができます。
ミオグロビンは典型的な筋肉タンパク質です(しかし、それは筋肉だけに見られるわけではありません)。
ミオグロビンは、大量に存在するマッコウクジラから抽出され、精製されます。
クジラ類は人間のような呼吸をします。肺を持っていると、呼吸過程を通じて空気を吸収しなければなりません。マッコウクジラは、筋肉に存在するミオグロビンに結合することで酸素を蓄積できる筋肉にできるだけ多くの酸素を取り込む必要があります。その後、クジラ類の代謝には酸素が必要なため、酸素はゆっくりと放出されます。マッコウクジラが吸収できる酸素の量と、ダイビング中に利用できる酸素の量が増えます。
ミオグリビンは可逆的に酸素と結合し、末梢組織に存在する割合が高くなります。これは、組織が時間的に離れた酸素供給を処理することに慣れているほどです。
<---ミオグロビンは筋肉に存在するタンパク質であり、その機能はまさに酸素の「貯蔵庫」の機能です。

肉を多かれ少なかれ赤くするのは、ヘムタンパク質の含有量です(肉を赤くするのはヘムです)。
ヘモグロビンはミオグロビンと多くの構造的類似性があり、可逆的な方法で分子状酸素に結合することができます。しかし、ミオグロビンは一般に筋肉や末梢組織に限定されていますが、ヘモグロビンは血液の40%を占める赤血球や赤血球(偽細胞、つまり実際の細胞ではありません)に見られます。
ミオグロビンとは対照的に、ヘモグロビンの働きは、肺で酸素を取り、それを必要な細胞に放出し、二酸化炭素を取り、そしてサイクルが再び始まる肺に放出することです。
L」ヘモグロビン これはテトラメーターです。つまり、それぞれがヘムグループを持ち、2つずつ同一である4つのポリペプチド鎖で構成されています(人間には2つのアルファ鎖と2つのベータ鎖があります)。
ヘモグロビンの主な機能は酸素の輸送です。ヘモグロビンが関与する血液の別の機能は、組織への物質の輸送です。
肺(酸素が豊富)から組織への経路では、ヘモグロビンは酸素を運びます(同時に他の物質が組織に到達します)が、逆の経路では、組織によって収集された廃棄物、特に炭素を運びます代謝で生成される二酸化炭素。
人間の発達には、特定の期間だけ発現する遺伝子があります。このため、成人男性の胎児、胚、さまざまなヘモグロビンがあります。
これらの異なるヘモグロビンを構成する鎖は異なる構造を持っていますが、実際にはいくつかの類似点があり、それらが実行する機能は多かれ少なかれ同じです。
いくつかの異なる鎖の存在の説明は次のとおりです。生物の進化過程の過程で、ヘモグロビンでさえ、酸素が豊富な領域から不足している領域への酸素の輸送に特化して進化しました。進化の連鎖の1 "ヘモグロビンは小さな生物で酸素を輸送しました;進化の過程で生物はより大きな次元に達したので、ヘモグロビンはそれが豊富な点からさらに離れた領域に酸素を輸送できるように変更されました;これを行うと、進化の過程で、ヘモグロビンを構成する鎖の新しい構造がコード化されます。
ミオグロビンは適度な圧力でも酸素と結合します;末梢組織には約30mmHgの圧力(PO2)があります:この圧力のミオグロビンは酸素を放出しないので、酸素運搬体としては効果がありません。一方、ヘモグロビンは、それはより弾力性のある振る舞いをします:それは酸素を高圧に結合し、圧力が下がるとそれを解放します。
タンパク質が機能的に活性である場合、その形状は少し変化する可能性があります。たとえば、酸素化ミオグロビンは非酸素化ミオグロビンとは形状が異なり、この変異は隣接するタンパク質に影響を与えません。
ヘモグロビンなどの関連タンパク質の場合は状況が異なります。鎖が酸素化すると、その形状が変化するように誘導されますが、この変更は3次元であるため、テトラメーターの他の鎖も影響を受けます。鎖が関連しているという事実は、1つの変更が、たとえ程度が異なっていても、他の隣接するものに影響を与えることを示唆しています。チェーンが酸素化すると、テトラメーターの他のチェーンは、酸素に対して「敵意の少ない態度」を取ります。それに近い鎖が順番に酸素化するにつれて、酸素化物は減少します。同じことが脱酸素にも当てはまります。
デオキシヘモグロビンの四次構造はT(時制)型と呼ばれ、オキシヘモグロビンの四次構造はR(放出)型と呼ばれます。緊張状態では、酸性アミノ酸と塩基性アミノ酸の間に一連のかなり強い静電相互作用があり、デオキシヘモグロビンの堅固な構造につながります(これが「緊張形態」である理由です)が、酸素が結合すると、これらの実体が相互作用が減少します(したがって、「解放された形式」)。さらに、酸素がない場合、ヒスチジンの電荷(構造を参照)はアスパラギン酸の反対の電荷によって安定化されますが、酸素が存在する場合、タンパク質の側でプロトンを失う傾向があります。これはすべて、酸素化ヘモグロビンが脱酸素化ヘモグロビンよりも強い酸であることを含みます。 ボーア効果.
pHに応じて、ヘム基は多かれ少なかれ酸素に結合しやすくなります。酸性環境では、ヘモグロビンは酸素を放出しやすくなります(緊張状態は安定します)が、塩基性環境では、酸素との結合はより困難になります。

各ヘモグロビンは、流入する酸素(O2)1モルあたり0.7プロトンを放出します。
ボーア効果により、ヘモグロビンは酸素を運ぶ能力を向上させることができます。
肺から組織に移動するヘモグロビンは、圧力、pH、温度の関数としてバランスをとる必要があります。
温度の影響を見てみましょう。
肺胞の温度は外部温度よりも約1〜1.5°C低く、筋肉の温度は約36.5〜37°Cです。温度が上昇すると、飽和係数が(同じ圧力で)低下します。これは、運動エネルギーが増加し、解離が促進されるために発生します。

ヘモグロビンが酸素に結合する能力に影響を与える可能性のある他の要因があり、その1つは2,3ビスホスホグリセリン酸の濃度です。
2,3ビスホスホグリセリン酸は、赤血球に4〜5 mMの濃度で存在する代謝物です(生物の他の部分では、このような高濃度で存在しません)。
生理学的pHでは、2,3ビスホスホグリセリン酸が脱プロトン化され、5つの負電荷があります。ヘモグロビンの2つのベータ鎖は高濃度の正電荷を持っているため、これらの鎖の間に挟まれています。ベータ鎖と2,3ビスホスホグリセリン酸の間の静電相互作用は、システムに一定の剛性を与えます。酸素との親和性がほとんどない緊張した構造が得られます。酸素化中に、2,3ビスホスホグリセリン酸が排出されます。
赤血球では、「c」は、1,3ビスホスホグリセリン酸(代謝によって生成される)を2,3ビスホスホグリセリン酸に変換して4〜5 mMの濃度に達するため、ヘモグロビンが組織内の「酸素」を交換できる特別な装置です。
組織に到達したヘモグロビンは放出状態(酸素に結合)ですが、組織の近くではカルボキシル化されて緊張状態になります。この状態のタンパク質は、酸素と結合する傾向が少なくなります。放出された状態になるため、ヘモグロビンは組織に酸素を放出します。さらに、水と二酸化炭素の反応により、H +イオンが生成されるため、ボーア効果によりさらに酸素が生成されます。
二酸化炭素は原形質膜を通過して赤血球に拡散します。赤血球は血液の約40%を占めるため、組織から拡散する二酸化炭素の40%のみが赤血球に流入すると予想されます。実際、二酸化炭素を変換する酵素が含まれているため、二酸化炭素の90%が赤血球に流入します。炭酸では、赤血球中の二酸化炭素の定常濃度が低く、したがって侵入率が高くなります。

赤血球が組織に到達したときに発生する別の現象は次のとおりです。勾配によって、「HCO3-(二酸化炭素の誘導体)は」赤血球を離れ、負の電荷の出力のバランスをとるために、「塩化物の侵入」があります。浸透圧の増加を決定します:この変動のバランスをとるために、赤血球の膨張を引き起こす水の侵入もあります(HAMBURGER効果)。赤血球が肺肺胞に到達すると、反対の現象が発生します:赤血球の収縮(HALDANE効果)したがって、(肺に向けられた)静脈赤血球は動脈赤血球よりも丸みを帯びています。