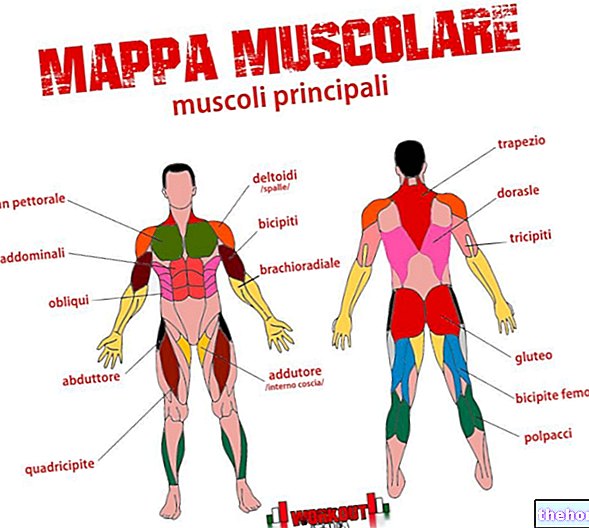
ミエリンは、主に脂質とタンパク質からなるラメラ構造の絶縁物質です。白灰色がかった光景で、麦わら色の色合いで、ミエリンはニューロンの軸索を外部から覆っています。このコーティングは、単純なもの(単層)にすることも、さまざまな同心円状の層で構成することもできます。これにより、一種のシースまたはスリーブが生成されます。
タンパク質
脂質
ガングリオシド
コレステロール
セレブロシド
セレブロシド硫酸塩(スルファチド)
ホスファチジルコリン(レシチン)
ホスファチジルエタノールアミン(セファリン)
ホスファチジルセリン
スフィンゴミエリン
その他の脂質
21.3
78.7
0.5
40.9
15.6
4.1
10.9
13.6
5.1
4.7
5.1
軸索を取り巻くミエリンの層に応じて、無髄神経線維(実際の鞘がない単層)と有髄神経線維(多層スリーブ)について話します。ミエリンがある場合、神経組織は白っぽく見えます。したがって、私たちは白質について話します。ミエリンがない場合、神経組織は灰色がかったように見えます。したがって、灰白質について話します。
中枢神経系では、軸索は一般に有髄ですが、末梢レベルでは、交感神経線維のほとんどの周りにミエリン鞘がありません。
後で見るように、ミエリン鞘の形成は、オリゴデンドロサイト(中枢神経系のミエリンの場合)およびシュワン細胞(末梢神経系のミエリンの場合)に委ねられています。ニューロンの軸索を取り囲むミエリンは、本質的にシュワン細胞(末梢神経系)とオリゴデンドロサイト(中枢神経系)の原形質膜で構成されています。
ミエリンの主な機能は、神経インパルスの正しい伝導を可能にし、いわゆる「跳躍伝導」を介してそれらの伝達速度を増幅することです。

ミエリンの二次的であるが等しく重要な機能は、それが覆う軸索に対する機械的保護と栄養維持の機能です。
代わりに、ミエリンがない場合、ニューロン(特にニューロンネットワークが特に密集しているCNSレベル)は興奮性であり、絶縁カバーのない電線と同じように、周囲の多くの信号に応答するため、絶縁機能は重要です。目的地に持ってこないで電流を分散させます。
ミエリンの組成を調べると、脂質、特にコレステロール、および程度は低いがレシチンやセファリンなどのリン脂質からの優勢な寄与に注目します。代わりに、タンパク質の80%は、塩基性タンパク質とプロテオリピドタンパク質で構成されています。マイナーなタンパク質もあり、その中でいわゆるオリゴデンドロサイトタンパク質が際立っています。


予想通り、ミエリンは特定の細胞の原形質膜(形質膜)で構成されており、軸索の周りを数回包み込んでいます。中枢神経系のレベルでは、ミエリンはオリゴデンドロサイトと呼ばれる細胞によって生成されますが、末梢レベルでは同じ機能がシュワン細胞によってカバーされています。どちらの細胞タイプもいわゆるグリア細胞に属しています。ミエリンは、これらのグリア細胞が細胞膜で軸索を包み込み、細胞質を外側に押し出して、各コイルが2つの層の追加に対応するときに形成されます。明確にするために、髄鞘形成プロセスは、収縮したバルーンを鉛筆の周りに巻き付けること、またはガーゼの二重層を指の周りに巻き付けることと比較することができます。
S.N.C.以来スペースの問題があり、各オリゴデンドロサイトは1つのセグメントにのみミエリンを提供しますが、より多くの軸索を提供します。したがって、各軸索は、異なるオリゴデンドロサイトによって形成された有髄セグメントに囲まれています。しかし、末梢レベルでは、個々のシュワン細胞がミエリンを単一の軸索に供給します。
オリゴデンドロサイトとシュワン細胞は、軸索の直径からミエリンを生成するように誘導されます。CNSでは直径が0.3μmのときにこれが発生しますが、SNPでは2μmを超える直径から始まります。

構造的に無髄の繊維は、裸の軸索の小さな束で構成されています。各束はシュワン細胞に包まれており、シュワン細胞は薄い細胞質の分枝を送って個々の軸索を分離します。したがって、無髄線維では、単一のシュワン細胞の内屈曲部に多数の小径軸索が含まれている可能性があります。
末梢レベルでは、シュワン細胞によって生成されたミエリンの存在が神経線維を再生する可能性を与えます。これは、数年前まで中枢神経系のレベルでは不可能であると考えられていました。実際、シュワン細胞とは異なり、オリゴデンドロサイトは損傷時に神経線維の再生を促進しません。しかし、最近の研究では、再生は困難であるが中枢神経系でも可能であり、潜在的に「神経発生」または新しいニューロンの形成さえ可能であることが示されています。