読書に戻る:肥大トレーニングルーチン
可用性は、L6筋細胞におけるホスファチジルイノシトール3-キナーゼ、p70 S6キナーゼおよびグリコーゲンシンターゼキナーゼ-3活性を調節します:システムAアミノのL-ロイシン誘発性アップレギュレーションにおけるラパマイシンの哺乳類標的(mTOR)経路の関与の証拠酸輸送KarinePEYROLLIER、Eric HAJDUCH、Anne S. BLAIR、Russell HYDE、Harinder S. HUNDAL1 メディカルサイエンスインスティテュート/ウェルカムトラストバイオセンターコンプレックス、ダンディー大学、ダンディーDD1 5EH、英国、ダウストリートの解剖生理学部ラパマイシン経路の哺乳類標的は、インスリン受容体基質1の細胞内再分布を介してインスリンシグナル伝達を調節し、インスリンの栄養シグナルと代謝シグナルを統合します 高野敦子、碓氷伊佐尾、春田哲郎*、川原淳子、宇野達仁、岩田稔、小林正史 富山医科薬科大学第一医学部、富山930-0194、日本 高い食事炭水化物制限の効果 タンパク質代謝および体細胞向性軸に関するタンパク質摂取量マシューP.ハーバー、サイモンシェンク、アリエルL.バーカン、ジェフリーF.ホロウィッツ運動学部(MPH、SS、JFH)および内分泌代謝学(ALB)、ミシガン大学および退役軍人Affairs Medical Center、Ann Arbor、Michigan 48109 ヒト骨格筋タンパク質合成に対するインスリンの効果は、筋肉の血流とアミノ酸の利用可能性におけるインスリン誘発性の変化によって調節されます。 Fujita S、Rasmussen BB、Cadenas JG、Grady JJ、Volpi E.南カリフォルニア大学医学部、ロサンゼルス、カリフォルニア、米国 血中アミノ酸の利用可能性が低下した場合、短期間のインスリンと栄養エネルギーの供給は筋肉タンパク質合成を刺激しません。Bell JA、Fujita S、Volpi E、Cadenas JG、Rasmussen BB Sealy Center on Aging&Stark Diabetes Center、Department of Physical Therapy、University of Texas Medical Branch、301 University Blvd.、Galveston、TX 77555-1144、USA 骨格筋におけるアミノ酸と抵抗運動によるmTORの調節。DeldicqueL、Theisen D、Francaux M. Institut d "Education Physique et de Redaptation、Universite catholique de Louvain、ベルギー インシュリンの筋肉への同化作用のメカニズム:アミノアシルtRNAおよび他の代理測定を使用した筋肉タンパク質合成および分解の測定。ChowLS、Albright RC、Bigelow ML、Toffolo G、Cobelli C、NairKS。内分泌学部門、メイヨークリニック医学部、200 First St. SW、ロチェスター、MN 55905、米国。回復後の筋肉タンパク質動態およびアミノ酸輸送に対するインスリン作用 レジスタンスエクササイズGianniBiolo、Bradley D.Williams、R.Y。デクラン・フレミング、ロバート・R・ウォル・フェ・スガA、平野T、影山H、大阪T、難波Y、辻M、三浦M、足立M、井上S 痩せたおよびVMH損傷肥満ラットの血漿レプチン、インスリン、およびインスリン抵抗性に対するフルクトースおよびグルコースの影響。 Am J Physiol Endocrinol Metab 2000 Apr; 278:E677-83 Romon M、Lebel P、Velly C、Marecaux N、Fruchart JC、DallongevilleJ。炭水化物または脂肪の食事に対するレプチンの反応とその後の満腹感およびエネルギー摂取量との関連。 Am J Physiol 1999 Nov; 277(5 Pt 1):E855-61 Hawkins M、Barzilai N、Liu R、Hu M、Chen W、RossettiL。脂肪誘発性インスリン抵抗性におけるグルコサミン経路の役割。 J Clin Invest 1997 May 1; 99:2173-82 Wang J、Liu R、Hawkins M、Barzilai N、Rossetti L.栄養素感知経路は、筋肉と脂肪のレプチン遺伝子発現を調節します。 Nature 1998 Jun 18; 393:684-8 Dear JF、Kolaczynski JW、Nyce MR、Ohannesian JP、Opentanova I、Goldman WH、Lynn RB、Zhang PL、Sinha MK、Considine RV肥満における脳脊髄液/血清レプチン比の低下:レプチン耐性のメカニズムの可能性。ランセット1996年7月20日; 348:159-61 Banks WA、Clever CM、FarrellCL。正常体重のマウスにおけるレプチンの血液から脳への輸送における部分的飽和および局所的変動。 Am J Physiol Endocrinol Metab 2000 Jun; 278:E1158-65 Banks WA、DiPalma CR、Farrell CL肥満の血液脳関門を通過するレプチンの輸送障害。ペプチド1999年11月; 20:1341-5 Kastin AJ、Akerstrom V.絶食は、副腎摘出術ではなく、脳へのレプチンの輸送を減少させます。ペプチド2000年5月; 21:679-82 Banks WA、DiPalma CR、Farrell CL肥満の血液脳関門を通過するレプチンの輸送障害。ペプチド1999年11月; 20:1341-5 J Endocrinol 2002 Jul; 174:121摂食状態でのTSH分泌の調節におけるレプチンの役割:invivoおよびinvitro研究 Ortiga-Carvalho TM、Oliveira KJ、Soares BA、Pazos-Moura CC Int J Mol Med 2002 Jan; 9:31 ラットの甲状腺は長い形のレプチン受容体を発現し、レプチンは甲状腺機能正常の絶食していない動物の腺の機能を刺激します Nowak KW、Kaczmarek P、Mackowiak P、Ziolkowska A、Albertin G、Ginda WJ、Trejter M、Nussdorfer GG、Malendowicz LK Clin Endocrinol Metab 20025月; 87:2391 低用量のレプチン投与は、エネルギー消費と甲状腺ホルモンの循環濃度に対する持続的な体重減少の影響を逆転させます ローゼンバウムM、マーフィーEM、ヘイムスフィールドSB、マシューズDE、レイベルRL J Endocrinol 2003 Jan; 176:7-12 TSHは脂肪細胞への直接効果によってレプチン分泌を刺激しますMenendezC、Baldelli R、Camina JP、Escudero B、Peino R、Dieguez C、Casanueva FF J Clin Endocrinol Metab 2001 Jul; 86:3284 レプチンおよびメラノコルチンシグナル伝達による甲状腺刺激ホルモン放出ホルモン遺伝子の転写調節。 Harris M、Aschkenasi C、Elias CF、Chandrankunnel A、Nillni EA、Bjoorbaek C、Elmquist JK、Flier JS、Hollenberg AN 内分泌学。 1998年5月; 139:2600-4。ラット視索上核および室傍核のオキシトシンおよびバソプレシンニューロンにおけるエストロゲン受容体ベータメッセンジャーリボ核酸の発現。 Hrabovszky E、Kallo I、Hajszan T、Shughrue PJ、Merchenthaler I、Liposits Z Jニューロビオール。 1998年9月5日; 36:357-78ラット脳全体のエストロゲン受容体(ERalphaおよびERbeta)の発現と神経ペプチド作動性の特徴:各サブタイプの異なる役割の解剖学的証拠。 Laflamme N、Nappi RE、Drolet G、Labrie C、Rivest S Acta Neurobiol Exp(Warsz)。 1996; 56:787-96。雌ラットにおける黄体形成ホルモン分泌の絶食誘発抑制を媒介する神経内分泌機構。前田K、塚村H 内分泌学。 2003年2月; 144:594-8。下垂体とは無関係にテストステロン分泌を調節する脳と精巣の間の神経経路の構成要素としての視床下部の室傍核の重要性。セルヴィッチDJ、リビエC Men1 Virve Luukkaa、Ullamari Pesonen、Ilpo Huhtaniemi、Aapo Lehtonen、Reijo Tilvis、Jaakko Tuomilehto、Markku Koulu、RistoHuupponenにおける血清テストステロンとレプチンの逆相関 食事の炭水化物は人間の栄養に不可欠ですか?エリックCウェストマン 人間の高強度の運動と筋肉グリコーゲンの利用可能性。 Balsom PD、Gaitanos GC、Soderlund K、Ekblom B 人間の長時間の運動中の筋肉代謝:炭水化物の利用可能性の影響。 McConell G、Snow RJ、Project J、Hargreaves M 長時間の疲労運動中の骨格筋エネルギー代謝。マークA. 2月とジェーンダンシー ケトジェニックダイエット、ライルマクドナルド(P 120) さまざまな強度の体重抵抗運動中の筋肉グリコーゲン分解。 R. A. Robergs、D。R。Pearson、D。L。Costill、W。J。Fink、D。D。Pascoe、M.A。ベネディクト、C。P。ランバートおよびJ. J.ザックウェイジャヒューマンパフォーマンスラボラトリー、ボール州立大学、マンシー、インディアナ47306 食後の運動における運動後のケトーシス:ヒトにおけるグルコースおよびアラニン摂取の影響。 J.H. Koeslag、L.I。 Levinrad、J.Dev。 Lochner 運動後のグリコーゲン合成の栄養と運動の決定要因。 Robergs RA
ホームページとボディリコンポジションメニュー-ボディリコンポジション
1.ボディリコンポジション-ボディリコンポジション 6.トレーニングにおけるエネルギーバランス 2.熱力学とトレーニング 7.レティーナとトレーニング 3.トレーニングにおけるタンパク質の重要性 8.トレーニングとインスリン感受性 4.トレーニングにおける糖質要件 9.肥大トレーニングルーチン 5.アントボリックダイエットと脂肪組織の設定値 10.リファレンス
タグ:
エクササイズ-ピラティス 外科的介入 腸の健康






エディターズチョイス
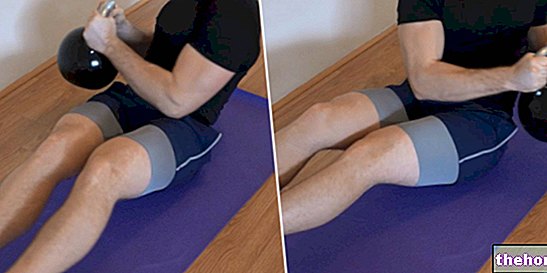 ロシアのひねり
ロシアのひねり
ビデオエクササイズ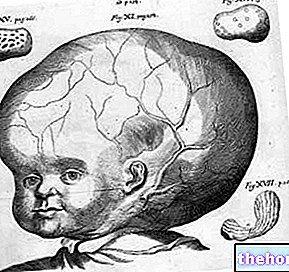 水頭症
水頭症
神経系の健康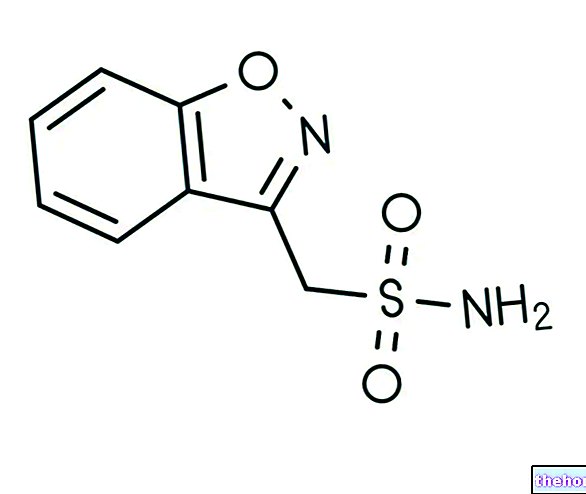 ゾニサミド:それが何であるか、それがいつどのように摂取されるか
ゾニサミド:それが何であるか、それがいつどのように摂取されるか
薬-a-cosa-serve.jpg) 眼底検査(検眼鏡):それは何のためですか?
眼底検査(検眼鏡):それは何のためですか?
目の健康 Janumet-添付文書
Janumet-添付文書
チラシ
最も読みやすい