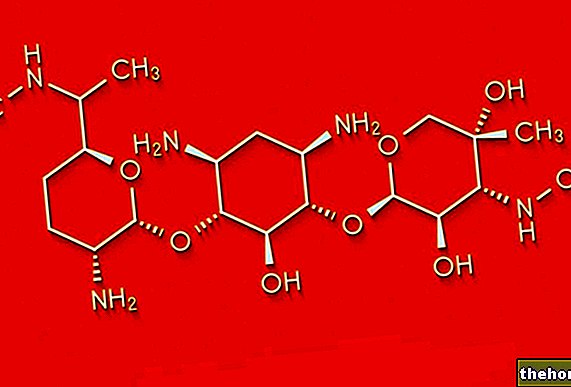
アソシエーション(または「連結」または「リンケージ」)
これまで、異なる対立遺伝子ペアが実際には異なる相同染色体ペアに見られると仮定して、メンデルのジハイブリッド(またはポリハイブリッド)交差を扱ってきましたが、染色体ペアの数は、種ごとに異なりますが、狭い範囲内で異なります(少数の種)遺伝子の数は数万に数えられますが、約100の染色体に達します)。
メンデルが実験のために選んだキャラクターがすべて独立して(したがって、ポリハイブリッドクロスのF2の表現型の分布の計算を混乱させることなく)分離したことは幸運なチャンスでした。隣接する遺伝子座で2対の対立遺伝子が見つかった場合、その法則は次のようになります。従うことは、連合の法則と呼ばれるでしょう。
非常に多くのキャラクターが単一の染色体上に遺伝子座を持ち、減数分裂で独立して分離する染色体ペアであることを知っているので、2つのキャラクターのペアが親の染色体に関連付けられている場合、どのくらいの頻度で発生するかをよく理解できます生物は、関連したままです。同様に、配偶子においても、したがって、それがそれ自身の遺伝物質をもたらす生物においても同様です。
したがって、メンデルの第3法則で発表された独立性に対する「関連性は、まれではない」例外を表すことがわかります。
交換または「乗換え」および組換え
減数分裂について言えば、遺伝物質の混合には2つの異なる瞬間があることを示しました。1つは配偶子の染色体分離の瞬間であり、もう1つはメンデルによって観察された瞬間です。
実際に先行するもう1つの瞬間は、相同染色体の各ペアの4つの染色分体が相互に同一の特性を交換する瞬間です。この交換に続いて、同じ染色体に関連付けられていた2つの因子が配偶子で独立します。確率交換が行われることは、最初の近似、染色体の長さに比例し、より長い染色体では、交換よりもさらに多くのことがあり得ます。
この現象は、顕微鏡下で十分な数の減数分裂を観察することにより、細胞学的に検出することができます。
再一致率は、親世代で関連付けられていた2つの文字がF2で異なる方法で再結合する率です。
2つの遺伝子座が完全に隣接している場合、キアズムがそれらを分離する確率は実質的にゼロになります。組換え率は次のようになります:n°組換え体。 2つの遺伝子座が2つの異なる染色体上にある場合、組換え率は0.5になります(P世代で結合された2つのキャラクターが、F2でランダムに一緒に見つかる確率は同じです)。したがって、組換え率は0.0から0.5の間で変動する可能性があります。染色体上の距離が小さい場合、距離と組換え率は正比例します。長距離の場合、2つの遺伝子座間で2つの交換が行われる可能性があります。これで、2つの交換によって分離された2つの要因が再び関連付けられていることが明らかになります。この時点で、遺伝子座の距離と組換えの確率との間の比例関係が失われていることは明らかです。
同じ染色体上に関連していることがわかった遺伝子座は、「関連グループ」を構成します。非常に離れた遺伝子座は、交換による分離の可能性が非常に高いため、独立しているように動作しますが、それぞれが、より低い組換え率で、中間遺伝子座に関連付けられます。
アソシエーショングループ内の遺伝子の多くのペア間の組換え率がわかっている場合、「遺伝地図」の構築を開始できます。 2つの遺伝子(aとb)間の距離は組換え率で表され、3番目の遺伝子cからのaの距離は、bからの距離に対する合計または差のいずれかである可能性があることに留意してください。相互距離のマップを再構築します。これは、その関連グループ内、つまりその染色体の遺伝子マップになります。
ここで、遺伝子型の特徴の表現型の発現を制限するいくつかの概念を一般的に検討する必要があります。
まず、浸透度と表現力の概念について説明し、次に遺伝子作用の調節現象に特に注意を向けます。
浸透度
遺伝子の浸透度は、表現型に現れる能力を表しています。浸透度は、浸透度を含む100の遺伝子型のうち、その文字を示す表現型の頻度を数えることによって統計的に測定されます。浸透度が0.7の形質は、その遺伝子型頻度の70%で表現型的に発生する形質です。
表現度
表現度は、表現型の発現の程度の定量的評価です。
遺伝子発現の調節
細胞はすべての酵素とタンパク質を同じ速度で同時に生成します。たとえば、大腸菌細胞は、ベータガラクトシダーゼ酵素のおかげでグルコースとガラクトースに分解できるため、ラクトース二糖からエネルギーと炭素原子を供給することができます。ラクトースを含む可能性のある通常の大腸菌では、ベータガラクトシダーゼの分子は約3000分子あり、その細胞のタンパク質の3%に相当します。ラクトースがない場合、細菌細胞ごとにベータガラクトシダーゼの分子は1つだけになります。ガラクトシダーゼは、使用可能になったときに新しいmRNA分子から合成されます。ラクトースが存在しない場合でも、酵素が豊富な大腸菌の変異株が知られています。これらの変異株は、基質なしで残る酵素を生成するためにエネルギーと材料を不必要に消費することを余儀なくされるため、正常細胞と比較して不利です。乳糖の場合のように、酵素の量を増加させる物質は誘導物質と呼ばれ、酵素は誘導性であると言われます。他の物質は、これらも特定の方法で、特定の酵素の生成を誘発します。また、例えば、炭素およびアンモニウム(NH3)を有するそのすべてのアミノ酸を構築することができるE.coliでは、特定のアミノ酸(例えば、ヒスチジン)の培養培地中の存在は、アミノ酸自体の生合成:これらの酵素は抑制可能であると言われます細菌細胞では、mRNA分子は形成後すぐに破壊されるため、mRNAの生成を制御することは同時に酵素合成を制御することを意味します時間。
オペロン
バクテリア細胞がどのように酵素のそれ自身の生産を制御することができるかを説明するために、ジェイコブとモノーはオペロンの仮説を定式化しました。オペロンは、DNAのストレッチに沿って不連続性なしに機能的に関連し、整列しているいくつかの遺伝子によって形成されます。オペロンは、3つの異なるタイプの遺伝子で構成されています。制御が行われるオペレーター;酵素または他のタンパク質をコードする1つまたは複数の構造遺伝子。ベータガラクトシダーゼシステムでは、オペロンには、ベータガラクトシダーゼのオペロンに加えて、構造をコードする他の2つの遺伝子も含まれます。ラクトースの代謝に関与する他の酵素。これらの遺伝子は互いに隣接しており、同じDNAらせんに沿って次々と転写されて単一のmRNA分子になります。こうして生成されたmRNA分子は非常に短時間活性であり、その後特定の酵素によって破壊されます。
オペロンの活動は、別の遺伝子であるレギュレーターによって制御されます。レギュレーターは、オペロンから離れている場合もあります。このレギュレーターは、リプレッサーと呼ばれるタンパク質をコードしており、オペレーター遺伝子のDNAに結合しているように見えます。プロモーターと構造遺伝子の間は実際にmRNAの生成をブロックします。
次に、リプレッサーが制御され、制御は「信号」物質によって実行されます。誘導性酵素の場合、この物質は「インデューサーです。インデューサー」はリプレッサー分子に結合してその形状を変更し、DNAに適応できなくなります。この場合、プロモーターと構造遺伝子の間にリプレッサーがないためです。 、リプレッサーはmRNA分子を形成し、これらからタンパク質分子を形成することができます。インデューサーの供給が再び枯渇すると、レギュレーターは制御を取り戻し、新しいmRNA、したがって新しいタンパク質の生成を停止します。ベータガラクトシダーゼシステムでは、インデューサーはラクトースまたはこれに非常に類似した物質です。酵素の生合成を可能にするためにそれを不活性化するリプレッサーに加わります。抑制可能な酵素の場合、「シグナル」として機能する物質はコリプレッサーとして機能します。リプレッサーは、コリプレッサーと組み合わされた場合にのみアクティブになります。十数種類の酵素が関与するヒスチジンシステムでは、このアミノ酸がそのtRNAであるコリプレッサーであるヒスチジンと組み合わされています。
アロステリック相互作用
酵素の形状を変えることによる酵素の不活性化を含むアロステリック相互作用は、細胞の代謝活性を調節する異なる方法を提供します。アロステリック相互作用は、オペロンのインダクター-リプレッサーシステムよりも正確な制御を可能にしますが、特定の物質の生合成を最初の段階から除外するという有用な結果、つまりmRNAの生成を達成しません。
EUCARIUSの制御システム
オペロンと同様の調節システムが機能しており、植物や動物の間で卓越していると信じるに至るいくつかの事実があります。これらの生物の染色体は、大腸菌や他の原核生物の染色体とは大きく異なります。これらの遺伝子の制御細胞は非常に異なります。有糸分裂のメカニズムは、特定の植物または動物の各細胞がすべての情報を持っているようなものです。
受精卵に存在する遺伝学したがって、特殊な細胞のほとんどの遺伝子は、細胞の寿命を通して非効率のままです。これらの細胞のDNAは常にタンパク質と関連しています。したがって、真核生物の遺伝子抑制には、正確にこの関連が必要である可能性があります。 DNAとタンパク質の間。